
The goal of my lab is to understand the hippocampus in terms of the menagerie of cell types it comprises. This requires a description of the cell types, in terms of gene expression, structure, and function, as well as determination of the synaptic connections between them and their outputs to other brain structures. The following provides some background about how I arrived at these interests and where it is taking us.
As an undergraduate at the University of British Columbia, I did research in a lab studying the neurophysiological basis of epilepsy. I made field potential recordings from hippocampal slices under conditions that caused them to generate rhythmic discharges, which are considered seizure-like in that many neurons generate synchronous discharges. I was fascinated by the notion that a tiny slice of brain tissue could generate spontaneous activity, causing me to wonder, “how does it work?”
My reading at the time included the works of many great neurophysiologists (e.g. Kandel, Llinás, Prince, Schwartzkroin), but I was drawn to the elegance of Dan Johnston’s work on the subject, which motivated me to do graduate research in his lab at the Baylor College of Medicine. While there, I became fascinated by the notion the neural network activity was dictated by the properties of the neurons in a circuit (including their structure and the ion channels that determined their excitable properties) as well as the synaptic connections among them.
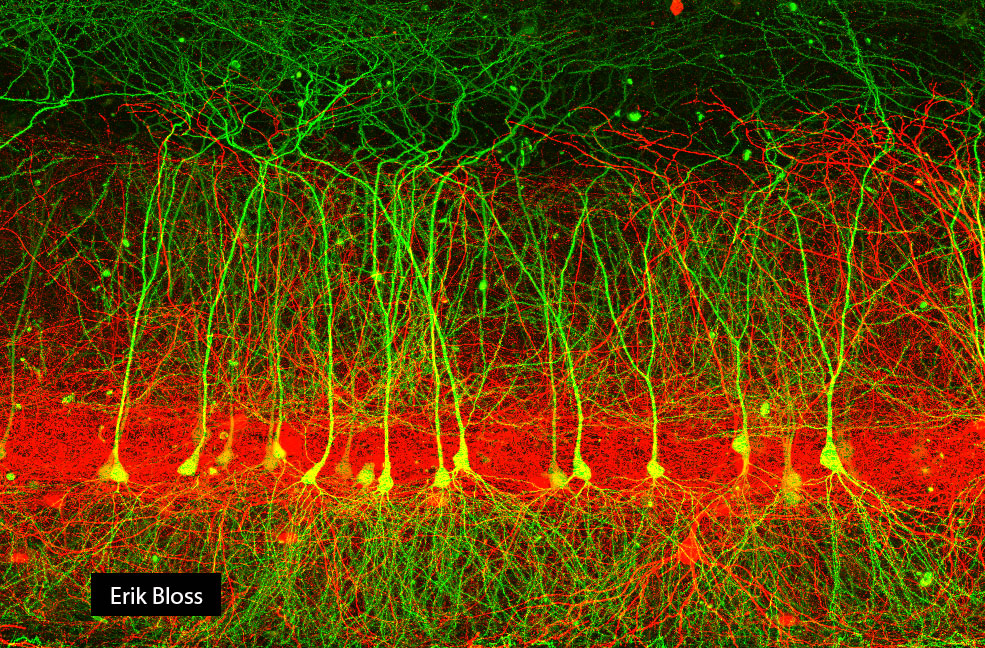
In the late 1980s and early 1990s, powerful methods were being applied to the study of neurons in slices. While in the Johnston lab, I used patch-clamp recordings to study the properties of multiple types of hippocampal neurons. At the time, it was apparent to me that these methods were being advanced to new levels in Bert Sakmann’s lab at the Max Planck Institute in Heidelberg, so I went there to continue my studies of hippocampal neurons as a postdoc.
One big advance in Sakmann’s lab at the time was that patch-clamp recordings in slices were being performed under visual control, making these experiments much easier than they were by recording almost blindly in the slice. Greg Stuart exploited and improved on these visually guided recordings to establish methods for recording from dendrites in slices. This was a breakthrough, because it meant that the dendritic tree—vast territory that was previously inaccessible to electrophysiological recording—suddenly became available for exploration and discovery.
Probing the cellular function of the nervous system, one quickly realizes that neurons are fascinating and complex machines. It seems inescapable to me that we won’t really understand how neural circuits function—whether that be during synchronous activity in a slice or during normal activity in a behaving animal—unless we determine the computational properties of individual neurons. Most neurons do not appear to operate like simple McCullough–Pitts integrate-and-fire units. Rather, they integrate their inputs in complex ways that are not yet completely understood.
Studying how one type of pyramidal neuron (CA1) integrates the tens of thousands of synaptic inputs it receives has consumed much of my time for decades, but I also realize that neural circuit function arises from the concerted operation of a host of neurons with complex morphologies and functional properties.
I initially became interested in the hippocampus because of its potential importance in epilepsy, but I quickly became fascinated by the structure for other reasons. The well-known role of the hippocampus in memory intrigued me (both in man and in rodents) and like others, I was able to exploit the highly organized, laminar structure of the hippocampus experimentally. Other aspects of the hippocampus that fascinate me are its role in spatial navigation and the property of “place fields” that govern the firing of many hippocampal neurons. Understanding how the properties and synaptic connectivity of hippocampal neurons give rise to place-cell firing seems like a difficult and intriguing problem, yet one that is likely to be solved in a satisfying way within my lifetime. Related observations, such as the precession of spiking relative to the hippocampal theta rhythm, which occurs in a place cell as a rat or a mouse advances through the cell’s place field, or the sequences of spiking that occur both within and across cycles of the theta rhythm, also seem within the reach of a detailed understanding. Problems such as the mechanisms responsible for sequence replay within sharp-wave ripples, seem even more challenging, but within reach, given the impressive pace of progress in the field.
My own expertise in patch-clamp recording and computational modeling has allowed me to make some contributions to understanding how hippocampal neurons work, most notably CA1 pyramidal neurons in the hippocampus. None of these contributions were made on my own. I am fortunate to have worked with many talented, wonderful people over the years (see the gallery below and another on my Lab Alumni page). Now, at Janelia I have the good fortune of working with more talented people, not just in my own lab, but also in other labs and in the shared resources.
Collectively, our goal is to continue to contribute to a better understanding of how the hippocampus works, by further defining and functionally dissecting the constellation of different cell types that comprise this fascinating circuit.
References
Buzsáki G, Moser EI. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci. 16:130-8, 2013.
Johnston D, Brown TH. Control theory applied to neural networks illuminates synaptic basis of interictal epileptiform activity. Adv. Neurol. 44:263-74, 1986.
O'Keefe J. Place units in the hippocampus of the freely moving rat. Exp. Neurol. 51:78-109, 1976.
O'Keefe J, Recce ML. Phase relationship between hippocampal place units and the EEG theta rhythm. Hippocampus. 3:317-30, 1993.
Skaggs WE, McNaughton BL, Wilson MA, Barnes CA. Theta phase precession in hippocampal neuronal populations and the compression of temporal sequences. Hippocampus 6:149-72, 1996.
Skaggs WE, McNaughton BL. Replay of neuronal firing sequences in rat hippocampus during sleep following spatial experience. Science 271:1870-3, 1996.
Spruston N, Johnston D. Perforated patch-clamp analysis of the passive membrane properties of three classes of hippocampal neurons. J. Neurophysiol. 67:508-29, 1992.
Stuart GJ, Dodt HU, Sakmann B. Patch-clamp recordings from the soma and dendrites of neurons in brain slices using infrared video microscopy. Pflügers Arch. 423:511-8, 1993.





















