Filter
Associated Lab
- Aguilera Castrejon Lab (4) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (61) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (104) Apply Betzig Lab filter
- Beyene Lab (10) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (51) Apply Branson Lab filter
- Card Lab (37) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (15) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dennis Lab (2) Apply Dennis Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (43) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (19) Apply Espinosa Medina Lab filter
- Feliciano Lab (12) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (17) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (55) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (26) Apply Hermundstad Lab filter
- Hess Lab (77) Apply Hess Lab filter
- Ilanges Lab (3) Apply Ilanges Lab filter
- Jayaraman Lab (44) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (3) Apply Koay Lab filter
- Lavis Lab (146) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (7) Apply Li Lab filter
- Lippincott-Schwartz Lab (108) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (3) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (60) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (7) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (40) Apply Pachitariu Lab filter
- Pastalkova Lab (6) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (49) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (39) Apply Romani Lab filter
- Rubin Lab (111) Apply Rubin Lab filter
- Saalfeld Lab (47) Apply Saalfeld Lab filter
- Satou Lab (3) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (53) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (3) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (62) Apply Spruston Lab filter
- Stern Lab (77) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (38) Apply Stringer Lab filter
- Svoboda Lab (132) Apply Svoboda Lab filter
- Tebo Lab (11) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (19) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (41) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (4) Apply Voigts Lab filter
- Wang (Meng) Lab (28) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (9) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (43) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (18) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- High Performance Computing (7) Apply High Performance Computing filter
- Integrative Imaging (21) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (37) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Mass Spectrometry (1) Apply Mass Spectrometry filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Project Technical Resources (54) Apply Project Technical Resources filter
- Quantitative Genomics (20) Apply Quantitative Genomics filter
- Scientific Computing (103) Apply Scientific Computing filter
- Stem Cell & Primary Culture (14) Apply Stem Cell & Primary Culture filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2026 (59) Apply 2026 filter
- 2025 (222) Apply 2025 filter
- 2024 (210) Apply 2024 filter
- 2023 (157) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2838 Janelia Publications
Showing 1501-1510 of 2838 resultsPhotoreceptors for visual perception, phototaxis or light avoidance are typically clustered in eyes or related structures such as the Bolwig organ of Drosophila larvae. Unexpectedly, we found that the class IV dendritic arborization neurons of Drosophila melanogaster larvae respond to ultraviolet, violet and blue light, and are major mediators of light avoidance, particularly at high intensities. These class IV dendritic arborization neurons, which are present in every body segment, have dendrites tiling the larval body wall nearly completely without redundancy. Dendritic illumination activates class IV dendritic arborization neurons. These novel photoreceptors use phototransduction machinery distinct from other photoreceptors in Drosophila and enable larvae to sense light exposure over their entire bodies and move out of danger.
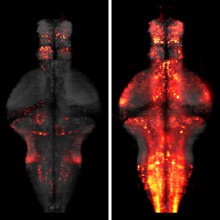
The processing of sensory input and the generation of behavior involves large networks of neurons, which necessitates new technology for recording from many neurons in behaving animals. In the larval zebrafish, light-sheet microscopy can be used to record the activity of almost all neurons in the brain simultaneously at single-cell resolution. Existing implementations, however, cannot be combined with visually driven behavior because the light sheet scans over the eye, interfering with presentation of controlled visual stimuli. Here we describe a system that overcomes the confounding eye stimulation through the use of two light sheets and combines whole-brain light-sheet imaging with virtual reality for fictively behaving larval zebrafish.

Developments in electrical and optical recording technology are scaling up the size of neuronal populations that can be monitored simultaneously. Light-sheet imaging is rapidly gaining traction as a method for optically interrogating activity in large networks and presents both opportunities and challenges for understanding circuit function.

The ability to visualize and quantitatively measure dynamic biological processes in vivo and at high spatiotemporal resolution is of fundamental importance to experimental investigations in developmental biology. Light-sheet microscopy is particularly well suited to providing such data, since it offers exceptionally high imaging speed and good spatial resolution while minimizing light-induced damage to the specimen. We review core principles and recent advances in light-sheet microscopy, with a focus on concepts and implementations relevant for applications in developmental biology. We discuss how light-sheet microcopy has helped advance our understanding of developmental processes from single-molecule to whole-organism studies, assess the potential for synergies with other state-of-the-art technologies, and introduce methods for computational image and data analysis. Finally, we explore the future trajectory of light-sheet microscopy, discuss key efforts to disseminate new light-sheet technology, and identify exciting opportunities for further advances.
The ways in which cells set the size of intracellular structures is an important but largely unsolved problem [1]. Early embryonic divisions pose special problems in this regard. Many checkpoints common in somatic cells are missing from these divisions, which are characterized by rapid reductions in cell size and short cell cycles [2]. Embryonic cells must therefore possess simple and robust mechanisms that allow the size of many of their intracellular structures to rapidly scale with cell size.
Binary cell fate decisions allow the production of distinct sister neurons from an intermediate precursor. Neurons are further diversified based on the birth order of intermediate precursors. Here we examined the interplay between binary cell fate and birth-order-dependent temporal fate in the Drosophila lateral antennal lobe (lAL) neuronal lineage. Single-cell mapping of the lAL lineage by twin-spot mosaic analysis with repressible cell markers (ts-MARCM) revealed that projection neurons (PNs) and local interneurons (LNs) are made in pairs through binary fate decisions. Forty-five types of PNs innervating distinct brain regions arise in a stereotyped sequence; however, the PNs with similar morphologies are not necessarily born in a contiguous window. The LNs are morphologically less diverse than the PNs, and the sequential morphogenetic changes in the two pairs occur independently. Sanpodo-dependent Notch activity promotes and patterns the LN fates. By contrast, Notch diversifies PN temporal fates in a Sanpodo-dispensable manner. These pleiotropic Notch actions underlie the differential temporal fate specification of twin neurons produced by common precursors within a lineage, possibly by modulating postmitotic neurons’ responses to Notch-independent transcriptional cascades.
Neurogenesis in Drosophila occurs in two phases, embryonic and post-embryonic, in which the same set of neuroblasts give rise to the distinct larval and adult nervous systems, respectively. Here, we identified the embryonic neuroblast origin of the adult neuronal lineages in the ventral nervous system via lineage-specific GAL4 lines and molecular markers. Our lineage mapping revealed that neurons born late in the embryonic phase show axonal morphology and transcription factor profiles that are similar to the neurons born post-embryonically from the same neuroblast. Moreover, we identified three thorax-specific neuroblasts not previously characterized and show that HOX genes confine them to the thoracic segments. Two of these, NB2-3 and NB3-4, generate leg motor neurons. The other neuroblast is novel and appears to have arisen recently during insect evolution. Our findings provide a comprehensive view of neurogenesis and show how proliferation of individual neuroblasts is dictated by temporal and spatial cues.
Most neurons of the central complex belong to 10 secondary (larvally produced) lineages. In the late larva, undifferentiated axon tracts of these lineages form a primordium in which all of the compartments of the central complex can be recognized as discrete entities. Four posterior lineages (DPMm1, DPMpm1, DPMpm2, and CM4) generate the classes of small-field neurons that interconnect the protocerebral bridge, fan-shaped body, noduli, and ellipsoid body. Three lineages located in the anterior brain, DALv2, BAmv1, and DALcl2, form the large-field neurons of the ellipsoid body and fan-shaped body, respectively. These lineages provide an input channel from the optic tubercle and connect the central complex with adjacent anterior brain compartments. Three lineages in the posterior cortex, CM3, CP2, and DPMpl2, connect the posterior brain neuropil with specific layers of the fan-shaped body. Even though all of the compartments of the central complex are prefigured in the late larval brain by the axon tracts of the above-mentioned lineages, the neuropil differentiates during the first 2 days of the pupal period when terminal branches and synapses of secondary neurons are formed. During this phase the initially straight horizontal layers of the central complex bend in the frontal plane, which produces the characteristic shape of the fan-shaped and ellipsoid body. Our analysis provides a comprehensive picture of the lineages that form the central complex, and will facilitate future studies that address the structure or function of the central complex at the single cell level.