Filter
Associated Lab
- Aso Lab (1) Apply Aso Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Clapham Lab (2) Apply Clapham Lab filter
- Fetter Lab (2) Apply Fetter Lab filter
- Harris Lab (7) Apply Harris Lab filter
- Hess Lab (8) Apply Hess Lab filter
- Ji Lab (11) Apply Ji Lab filter
- Lavis Lab (8) Apply Lavis Lab filter
- Lippincott-Schwartz Lab (6) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (7) Apply Liu (Zhe) Lab filter
- Magee Lab (2) Apply Magee Lab filter
- Rubin Lab (1) Apply Rubin Lab filter
- Saalfeld Lab (2) Apply Saalfeld Lab filter
- Schreiter Lab (1) Apply Schreiter Lab filter
- Shroff Lab (9) Apply Shroff Lab filter
- Singer Lab (1) Apply Singer Lab filter
- Svoboda Lab (2) Apply Svoboda Lab filter
- Tjian Lab (4) Apply Tjian Lab filter
- Turner Lab (1) Apply Turner Lab filter
Associated Project Team
Publication Date
- 2026 (1) Apply 2026 filter
- 2025 (4) Apply 2025 filter
- 2024 (2) Apply 2024 filter
- 2023 (4) Apply 2023 filter
- 2022 (3) Apply 2022 filter
- 2021 (2) Apply 2021 filter
- 2020 (4) Apply 2020 filter
- 2019 (7) Apply 2019 filter
- 2018 (6) Apply 2018 filter
- 2017 (8) Apply 2017 filter
- 2016 (12) Apply 2016 filter
- 2015 (11) Apply 2015 filter
- 2014 (8) Apply 2014 filter
- 2013 (4) Apply 2013 filter
- 2012 (5) Apply 2012 filter
- 2011 (7) Apply 2011 filter
- 2010 (3) Apply 2010 filter
- 2009 (2) Apply 2009 filter
- 2008 (8) Apply 2008 filter
- 2007 (2) Apply 2007 filter
- 2006 (1) Apply 2006 filter
- 2005 (1) Apply 2005 filter
- 1995 (1) Apply 1995 filter
- 1994 (2) Apply 1994 filter
- 1993 (2) Apply 1993 filter
- 1992 (4) Apply 1992 filter
- 1991 (2) Apply 1991 filter
Type of Publication
116 Publications
Showing 1-10 of 116 resultsTo image the accessible genome at nanometer scale in situ, we developed three-dimensional assay for transposase-accessible chromatin-photoactivated localization microscopy (3D ATAC-PALM) that integrates an assay for transposase-accessible chromatin with visualization, PALM super-resolution imaging and lattice light-sheet microscopy. Multiplexed with oligopaint DNA–fluorescence in situ hybridization (FISH), RNA–FISH and protein fluorescence, 3D ATAC-PALM connected microscopy and genomic data, revealing spatially segregated accessible chromatin domains (ACDs) that enclose active chromatin and transcribed genes. Using these methods to analyze genetically perturbed cells, we demonstrated that genome architectural protein CTCF prevents excessive clustering of accessible chromatin and decompacts ACDs. These results highlight 3D ATAC-PALM as a useful tool to probe the structure and organizing mechanism of the genome.
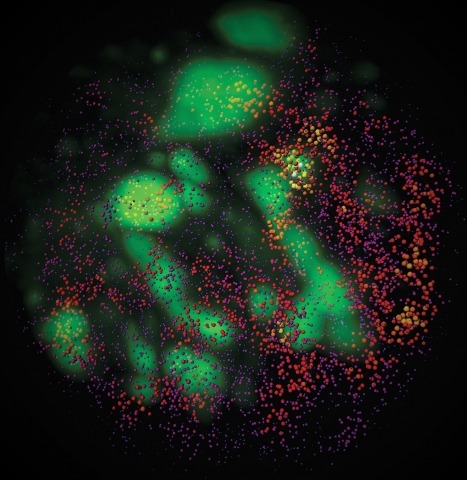
Combinatorial cis-regulatory networks encoded in animal genomes represent the foundational gene expression mechanism for directing cell-fate commitment and maintenance of cell identity by transcription factors (TFs). However, the 3D spatial organization of cis-elements and how such sub-nuclear structures influence TF activity remain poorly understood. Here, we combine lattice light-sheet imaging, single-molecule tracking, numerical simulations, and ChIP-exo mapping to localize and functionally probe Sox2 enhancer-organization in living embryonic stem cells. Sox2 enhancers form 3D-clusters that are segregated from heterochromatin but overlap with a subset of Pol II enriched regions. Sox2 searches for specific binding targets via a 3D-diffusion dominant mode when shuttling long-distances between clusters while chromatin-bound states predominate within individual clusters. Thus, enhancer clustering may reduce global search efficiency but enables rapid local fine-tuning of TF search parameters. Our results suggest an integrated model linking cis-element 3D spatial distribution to local-versus-global target search modalities essential for regulating eukaryotic gene transcription.
3D live imaging is important for a better understanding of biological processes, but it is challenging with current techniques such as spinning-disk confocal microscopy. Bessel beam plane illumination microscopy allows high-speed 3D live fluorescence imaging of living cellular and multicellular specimens with nearly isotropic spatial resolution, low photobleaching and low photodamage. Unlike conventional fluorescence imaging techniques that usually have a unique operation mode, Bessel plane illumination has several modes that offer different performance with different imaging metrics. To achieve optimal results from this technique, the appropriate operation mode needs to be selected and the experimental setting must be optimized for the specific application and associated sample properties. Here we explain the fundamental working principles of this technique, discuss the pros and cons of each operational mode and show through examples how to optimize experimental parameters. We also describe the procedures needed to construct, align and operate a Bessel beam plane illumination microscope by using our previously reported system as an example, and we list the necessary equipment to build such a microscope. Assuming all components are readily available, it would take a person skilled in optical instrumentation \~{}1 month to assemble and operate a microscope according to this protocol.
New methods in stem cell 3D organoid tissue culture, advanced imaging and big data image analytics now allow tissue scale 4D cell biology, but currently available analytical pipelines are inadequate for handing and analyzing the resulting gigabytes and terabytes of high-content imaging data. We expressed fluorescent protein fusions of clathrin and dynamin2 at endogenous levels in genome-edited human embryonic stem cells, which were differentiated into hESC-derived intestinal epithelial organoids. Lattice Light-Sheet Imaging with adaptive optics (AO-LLSM) allowed us to image large volumes of these organoids (70µm x 60µm x 40µm xyz) at 5.7s/frame. We developed an open source data analysis package termed pyLattice to process the resulting large (∼60Gb) movie datasets and to track clathrin-mediated endocytosis (CME) events. CME tracks could be recorded from ∼35 cells at a time, resulting in ∼4000 processed tracks per movie. Based on their localization in the organoid, we classified CME tracks into apical, lateral and basal events and found that CME dynamics are similar for all three classes, despite reported differences in membrane tension. pyLattice coupled with AO-LLSM makes possible quantitative, high temporal and spatial resolution analysis of subcellular events within tissues. Movie S1 Movie S1 Thresholded 3D AO-LLSM data of an intestinal epithelial organoid showing clathrin (red) and dynamin2 (green) puncta in surface depiction. The movie zooms out from a single clathrin mediated endocytosis event that shows both clathrin and dynamin2 at the same location to eventually show the whole AO-LLSM field of view. Nuclear envelopes and the outer membranes of the tissue are depicted in transparent white. Movie S2 Movie S2 Thresholded 3D AO-LLSM data of an intestinal epithelial organoid showing clathrin (red) and dynamin2 (green) puncta in surface depiction. The movie rotates the AO-LLSM field of view. Nuclear envelopes and the outer membranes of the tissue are depicted in transparent white. Movie S3 Movie S3 Thresholded 3D AO-LLSM data of an intestinal epithelial organoid. The curved surface is of the spherical organoid is visible as the movie rotates. Clathrin puncta are visible throughout the tissue (white). Movie S4 Movie S4 The detection step in the data processing pipeline retrieves all clathrin puncta in the volume. Detected puncta are marked with a cube (blue). Movie S5 Movie S5 Zoom on one clathrin puncta in the thresholded 3D dataset. The punctum of interest is marked with a blue cube. Other puncta are also visible. Movie S6 Movie S6 Zoom on the same clathrin puncta as in M3 in non-thresholded 3D data. The surrounding fluorescence is visible as a transparent cloud.
Imaging the 4D choreography of subcellular events in living multicellular organisms at high spatiotemporal resolution could reveal life’s fundamental principles. Yet extracting these principles from petabyte-scale image data requires fusing advanced light microscopy and cutting-edge machine learning models with biological insight and expertise.
Drosophila melanogaster has a rich repertoire of innate and learned behaviors. Its 100,000-neuron brain is a large but tractable target for comprehensive neural circuit mapping. Only electron microscopy (EM) enables complete, unbiased mapping of synaptic connectivity; however, the fly brain is too large for conventional EM. We developed a custom high-throughput EM platform and imaged the entire brain of an adult female fly at synaptic resolution. To validate the dataset, we traced brain-spanning circuitry involving the mushroom body (MB), which has been extensively studied for its role in learning. All inputs to Kenyon cells (KCs), the intrinsic neurons of the MB, were mapped, revealing a previously unknown cell type, postsynaptic partners of KC dendrites, and unexpected clustering of olfactory projection neurons. These reconstructions show that this freely available EM volume supports mapping of brain-spanning circuits, which will significantly accelerate Drosophila neuroscience..
How adherent and contractile systems coordinate to promote cell shape changes is unclear. Here, we define a counterbalanced adhesion/contraction model for cell shape control. Live-cell microscopy data showed a crucial role for a contractile meshwork at the top of the cell, which is composed of actin arcs and myosin IIA filaments. The contractile actin meshwork is organized like muscle sarcomeres, with repeating myosin II filaments separated by the actin bundling protein α-actinin, and is mechanically coupled to noncontractile dorsal actin fibers that run from top to bottom in the cell. When the meshwork contracts, it pulls the dorsal fibers away from the substrate. This pulling force is counterbalanced by the dorsal fibers' attachment to focal adhesions, causing the fibers to bend downward and flattening the cell. This model is likely to be relevant for understanding how cells configure themselves to complex surfaces, protrude into tight spaces, and generate three-dimensional forces on the growth substrate under both healthy and diseased conditions.
Developmental signals such as Wnts are often presented to cells in an oriented manner. To examine the consequences of local Wnt signaling, we immobilized Wnt proteins on beads and introduced them to embryonic stem cells in culture. At the single-cell level, the Wnt-bead induced asymmetric distribution of Wnt-β-catenin signaling components, oriented the plane of mitotic division, and directed asymmetric inheritance of centrosomes. Before cytokinesis was completed, the Wnt-proximal daughter cell expressed high levels of nuclear β-catenin and pluripotency genes, whereas the distal daughter cell acquired hallmarks of differentiation. We suggest that a spatially restricted Wnt signal induces an oriented cell division that generates distinct cell fates at predictable positions relative to the Wnt source.
Understanding biological systems requires observing features and processes across vast spatial and temporal scales, spanning nanometers to centimeters and milliseconds to days, often using multiple imaging modalities within complex native microenvironments. Yet, achieving this comprehensive view is challenging because microscopes optimized for specific tasks typically lack versatility due to inherent optical and sample handling trade-offs, and frequently suffer performance degradation from sample-induced optical aberrations in multicellular contexts. Here, we present MOSAIC, a reconfigurable microscope that integrates multiple advanced imaging techniques including light-sheet, label-free, super-resolution, and multi-photon, all equipped with adaptive optics. MOSAIC enables non-invasive imaging of subcellular dynamics in both cultured cells and live multicellular organisms, nanoscale mapping of molecular architectures across millimeter-scale expanded tissues, and structural/functional neural imaging within live mice. MOSAIC facilitates correlative studies across biological scales within the same specimen, providing an integrated platform for broad biological investigation. Preprint: https://www.biorxiv.org/content/early/2025/06/13/2025.06.02.657494
Macropinocytosis is a fundamental mechanism that allows cells to take up extracellular liquid into large vesicles. It critically depends on the formation of a ring of protrusive actin beneath the plasma membrane, which develops into the macropinocytic cup. We show that macropinocytic cups in Dictyostelium are organised around coincident intense patches of PIP3, active Ras and active Rac. These signalling patches are invariably associated with a ring of active SCAR/WAVE at their periphery, as are all examined structures based on PIP3 patches, including phagocytic cups and basal waves. Patch formation does not depend on the enclosing F-actin ring, and patches become enlarged when the RasGAP NF1 is mutated, showing that Ras plays an instructive role. New macropinocytic cups predominantly form by splitting from existing ones. We propose that cup-shaped plasma membrane structures form from self-organizing patches of active Ras/PIP3, which recruit a ring of actin nucleators to their periphery.