Filter
Associated Lab
- Aguilera Castrejon Lab (19) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (73) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Beyene Lab (15) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (55) Apply Branson Lab filter
- Card Lab (43) Apply Card Lab filter
- Cardona Lab (64) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (16) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (3) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (56) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (23) Apply Espinosa Medina Lab filter
- Feliciano Lab (12) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (30) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (65) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (30) Apply Hermundstad Lab filter
- Hess Lab (80) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (48) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (6) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (15) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (76) Apply Keller Lab filter
- Koay Lab (20) Apply Koay Lab filter
- Lavis Lab (158) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (32) Apply Li Lab filter
- Lippincott-Schwartz Lab (180) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (8) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (65) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (8) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (54) Apply Pachitariu Lab filter
- Pastalkova Lab (19) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (55) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (51) Apply Romani Lab filter
- Rubin Lab (149) Apply Rubin Lab filter
- Saalfeld Lab (64) Apply Saalfeld Lab filter
- Satou Lab (18) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (70) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (23) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (98) Apply Spruston Lab filter
- Stern Lab (160) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (43) Apply Stringer Lab filter
- Svoboda Lab (136) Apply Svoboda Lab filter
- Tebo Lab (35) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (53) Apply Turaga Lab filter
- Turner Lab (38) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (4) Apply Voigts Lab filter
- Wang (Meng) Lab (29) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (9) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2026 (74) Apply 2026 filter
- 2025 (222) Apply 2025 filter
- 2024 (210) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (192) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4269 Publications
Showing 3341-3350 of 4269 resultsThe mammalian retina conveys the vast majority of information about visual stimuli to two brain regions: the dorsal lateral geniculate nucleus (dLGN) and the superior colliculus (SC). The degree to which retinal ganglion cells (RGCs) send similar or distinct information to the two areas remains unclear despite the important constraints that different patterns of RGC input place on downstream visual processing. To resolve this ambiguity we injected a glycoprotein-deficient rabies virus coding for the expression of a fluorescent protein into the dLGN or SC; rabies virus labeled a smaller fraction of RGCs than lipophilic dyes like DiI but, crucially, did not label RGC axons of passage. ~80% of the RGCs infected by rabies virus injected into the dLGN were co-labeled with DiI injected into the SC, suggesting that many dLGN-projecting RGCs also project to the SC. However, functional characterization of RGCs revealed that the SC receives input from several classes of RGCs that largely avoid the dLGN - in particular, RGCs in which (1) sustained changes in light intensity elicit transient changes in firing rate and/or (2) a small range of stimulus sizes or temporal fluctuations in light intensity elicit robust activity. Taken together, our results illustrate several unexpected asymmetries in the information that the mouse retina conveys to two major downstream targets and suggest that differences in the output of dLGN and SC neurons reflect, at least in part, differences in the functional properties of RGCs that innervate the SC but not the dLGN.
The neocortex contains a multitude of cell types that are segregated into layers and functionally distinct areas. To investigate the diversity of cell types across the mouse neocortex, here we analysed 23,822 cells from two areas at distant poles of the mouse neocortex: the primary visual cortex and the anterior lateral motor cortex. We define 133 transcriptomic cell types by deep, single-cell RNA sequencing. Nearly all types of GABA (γ-aminobutyric acid)-containing neurons are shared across both areas, whereas most types of glutamatergic neurons were found in one of the two areas. By combining single-cell RNA sequencing and retrograde labelling, we match transcriptomic types of glutamatergic neurons to their long-range projection specificity. Our study establishes a combined transcriptomic and projectional taxonomy of cortical cell types from functionally distinct areas of the adult mouse cortex.
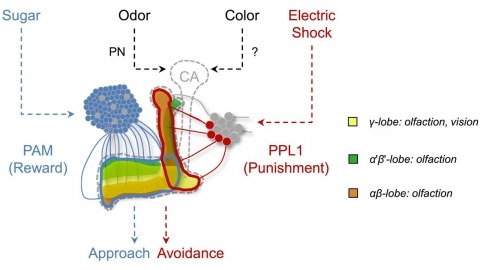
In nature, animals form memories associating reward or punishment with stimuli from different sensory modalities, such as smells and colors. It is unclear, however, how distinct sensory memories are processed in the brain. We established appetitive and aversive visual learning assays for Drosophila that are comparable to the widely used olfactory learning assays. These assays share critical features, such as reinforcing stimuli (sugar reward and electric shock punishment), and allow direct comparison of the cellular requirements for visual and olfactory memories. We found that the same subsets of dopamine neurons drive formation of both sensory memories. Furthermore, distinct yet partially overlapping subsets of mushroom body intrinsic neurons are required for visual and olfactory memories. Thus, our results suggest that distinct sensory memories are processed in a common brain center. Such centralization of related brain functions is an economical design that avoids the repetition of similar circuit motifs.

Light sheet-based fluorescence microscopy (LSFM) is emerging as a powerful imaging technique for the life sciences. LSFM provides an exceptionally high imaging speed, high signal-to-noise ratio, low level of photo-bleaching and good optical penetration depth. This unique combination of capabilities makes light sheet-based microscopes highly suitable for live imaging applications. There is an outstanding potential in applying this technology to the quantitative study of embryonic development. Here, we provide an overview of the different basic implementations of LSFM, review recent technical advances in the field and highlight applications in the context of embryonic development. We conclude with a discussion of promising future directions.
Historically, developmental-stage- and tissue-specific patterns of gene expression were assumed to be determined primarily by DNA regulatory sequences and their associated activators, while the general transcription machinery including core promoter recognition complexes, coactivators, and chromatin modifiers was held to be invariant. New evidence suggests that significant changes in these general transcription factors including TFIID, BAF, and Mediator may facilitate global changes in cell-type-specific transcription.
Previous studies showed that Roundabout (Robo) in Drosophila is a repulsive axon guidance receptor that binds to Slit, a repellent secreted by midline glia. In robo mutants, growth cones cross and recross the midline, while, in slit mutants, growth cones enter the midline but fail to leave it. This difference suggests that Slit must have more than one receptor controlling midline guidance. In the absence of Robo, some other Slit receptor ensures that growth cones do not stay at the midline, even though they cross and recross it. Here we show that the Drosophila genome encodes three Robo receptors and that Robo and Robo2 have distinct functions, which together control repulsive axon guidance at the midline. The robo,robo2 double mutant is largely identical to slit.
Slit is secreted by midline glia in Drosophila and functions as a short-range repellent to control midline crossing. Although most Slit stays near the midline, some diffuses laterally, functioning as a long-range chemorepellent. Here we show that a combinatorial code of Robo receptors controls lateral position in the CNS by responding to this presumptive Slit gradient. Medial axons express only Robo, intermediate axons express Robo3 and Robo, while lateral axons express Robo2, Robo3, and Robo. Removal of robo2 or robo3 causes lateral axons to extend medially; ectopic expression of Robo2 or Robo3 on medial axons drives them laterally. Precise topography of longitudinal pathways appears to be controlled by a combination of long-range guidance (the Robo code determining region) and short-range guidance (discrete local cues determining specific location within a region).
Rodent hippocampus exhibits strikingly different regimes of population activity in different behavioral states. During locomotion, hippocampal activity oscillates at theta frequency (5-12 Hz) and cells fire at specific locations in the environment, the place fields. As the animal runs through a place field, spikes are emitted at progressively earlier phases of the theta cycles. During immobility, hippocampus exhibits sharp irregular bursts of activity, with occasional rapid orderly activation of place cells expressing a possible trajectory of the animal. The mechanisms underlying this rich repertoire of dynamics are still unclear. We developed a novel recurrent network model that accounts for the observed phenomena. We assume that the network stores a map of the environment in its recurrent connections, which are endowed with short-term synaptic depression. We show that the network dynamics exhibits two different regimes that are similar to the experimentally observed population activity states in the hippocampus. The operating regime can be solely controlled by external inputs. Our results suggest that short-term synaptic plasticity is a potential mechanism contributing to shape the population activity in hippocampus.
BACKGROUND: Diastolic dysfunction is a poorly understood but clinically pervasive syndrome that is characterized by increased diastolic stiffness. Titin is the main determinant of cellular passive stiffness. However, the physiological role that the tandem immunoglobulin (Ig) segment of titin plays in stiffness generation and whether shortening this segment is sufficient to cause diastolic dysfunction need to be established. METHODS AND RESULTS: We generated a mouse model in which 9 Ig-like domains (Ig3-Ig11) were deleted from the proximal tandem Ig segment of the spring region of titin (IG KO). Exon microarray analysis revealed no adaptations in titin splicing, whereas novel phospho-specific antibodies did not detect changes in titin phosphorylation. Passive myocyte stiffness was increased in the IG KO, and immunoelectron microscopy revealed increased extension of the remaining titin spring segments as the sole likely underlying mechanism. Diastolic stiffness was increased at the tissue and organ levels, with no consistent changes in extracellular matrix composition or extracellular matrix-based passive stiffness, supporting a titin-based mechanism for in vivo diastolic dysfunction. Additionally, IG KO mice have a reduced exercise tolerance, a phenotype often associated with diastolic dysfunction. CONCLUSIONS: Increased titin-based passive stiffness is sufficient to cause diastolic dysfunction with exercise intolerance.
Large axon-diameter descending neurons are metabolically costly but transmit information rapidly from sensory neurons in the brain to motor neurons in the nerve cord. They have thus endured as a common feature of escape circuits in many animal species where speed is paramount. Though often considered isolated command neurons triggering fast-reaction-time, all-or-none escape responses, giant neurons are just one of multiple parallel pathways enabling selection between behavioral alternatives. Such degeneracy among escape circuits makes it unclear if and how giant neurons benefit prey fitness. Here we competed Drosophila melanogaster flies with genetically-silenced Giant Fibers (GFs) against flies with functional GFs in an arena with wild-caught damselfly predators and find that GF silencing decreases prey survival. Kinematic analysis of damselfly attack trajectories shows that decreased prey survival fitness results from GF-silenced flies failing to escape during predator attack speeds and approach distances that would normally elicit successful escapes. When challenged with a virtual looming predator, fly GFs promote survival by enforcing selection of a short-duration takeoff sequence as opposed to reducing reaction time. Our findings support a role for the GFs in promoting prey survival by influencing action selection as a means to enhance escape performance during realistically complex predation scenarios. Preprint: https://www.biorxiv.org/content/early/2024/05/01/2024.04.30.591368