Filter
Associated Lab
- Aguilera Castrejon Lab (16) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (64) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (112) Apply Betzig Lab filter
- Beyene Lab (13) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (52) Apply Branson Lab filter
- Card Lab (40) Apply Card Lab filter
- Cardona Lab (63) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (50) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (19) Apply Espinosa Medina Lab filter
- Feliciano Lab (7) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- Fitzgerald Lab (29) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (38) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (60) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (26) Apply Hermundstad Lab filter
- Hess Lab (76) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (46) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (6) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (76) Apply Keller Lab filter
- Koay Lab (18) Apply Koay Lab filter
- Lavis Lab (148) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (27) Apply Li Lab filter
- Lippincott-Schwartz Lab (167) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (6) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (61) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (46) Apply Pachitariu Lab filter
- Pastalkova Lab (18) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (51) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (43) Apply Romani Lab filter
- Rubin Lab (143) Apply Rubin Lab filter
- Saalfeld Lab (62) Apply Saalfeld Lab filter
- Satou Lab (16) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (67) Apply Schreiter Lab filter
- Sgro Lab (20) Apply Sgro Lab filter
- Shroff Lab (29) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (93) Apply Spruston Lab filter
- Stern Lab (156) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (33) Apply Stringer Lab filter
- Svoboda Lab (135) Apply Svoboda Lab filter
- Tebo Lab (33) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (21) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (49) Apply Turaga Lab filter
- Turner Lab (37) Apply Turner Lab filter
- Vale Lab (7) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (17) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (2) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (10) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (53) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (45) Apply GENIE filter
- Integrative Imaging (2) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (26) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2025 (72) Apply 2025 filter
- 2024 (223) Apply 2024 filter
- 2023 (163) Apply 2023 filter
- 2022 (193) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4064 Publications
Showing 4001-4010 of 4064 resultsPolarized fluorescence microscopy is a valuable tool for measuring molecular orientations in biological samples, but techniques for recovering three-dimensional orientations and positions of fluorescent ensembles are limited. We report a polarized dual-view light-sheet system for determining the diffraction-limited three-dimensional distribution of the orientations and positions of ensembles of fluorescent dipoles that label biological structures. We share a set of visualization, histogram, and profiling tools for interpreting these positions and orientations. We model the distributions based on the polarization-dependent efficiency of excitation and detection of emitted fluorescence, using coarse-grained representations we call orientation distribution functions (ODFs). We apply ODFs to create physics-informed models of image formation with spatio-angular point-spread and transfer functions. We use theory and experiment to conclude that light-sheet tilting is a necessary part of our design for recovering all three-dimensional orientations. We use our system to extend known two-dimensional results to three dimensions in FM1-43-labeled giant unilamellar vesicles, fast-scarlet-labeled cellulose in xylem cells, and phalloidin-labeled actin in U2OS cells. Additionally, we observe phalloidin-labeled actin in mouse fibroblasts grown on grids of labeled nanowires and identify correlations between local actin alignment and global cell-scale orientation, indicating cellular coordination across length scales.
Two-photon laser scanning microscopy of calcium dynamics using fluorescent indicators is a widely used imaging method for large-scale recording of neural activity in vivo. Here, we introduce volumetric two-photon imaging of neurons using stereoscopy (vTwINS), a volumetric calcium imaging method that uses an elongated, V-shaped point spread function to image a 3D brain volume. Single neurons project to spatially displaced 'image pairs' in the resulting 2D image, and the separation distance between projections is proportional to depth in the volume. To demix the fluorescence time series of individual neurons, we introduce a modified orthogonal matching pursuit algorithm that also infers source locations within the 3D volume. We illustrated vTwINS by imaging neural population activity in the mouse primary visual cortex and hippocampus. Our results demonstrated that vTwINS provides an effective method for volumetric two-photon calcium imaging that increases the number of neurons recorded while maintaining a high frame rate.
The hexameric AAA ATPase VPS4 facilitates ESCRT III filament disassembly on diverse intracellular membranes. ESCRT III components and VPS4 have been localized to the ciliary transition zone and spindle poles and reported to affect centrosome duplication and spindle pole stability. How the canonical ESCRT pathway could mediate these events is unclear. We studied the association of VPS4 with centrosomes and found that GFP-VPS4 was a dynamic component of both mother and daughter centrioles. A mutant, VPS4, which can't hydrolyze ATP, was less dynamic and accumulated at centrosomes. Centrosome localization of the VPS4mutant, caused reduced γ-tubulin levels at centrosomes and consequently decreased microtubule growth and altered centrosome positioning. In addition, preventing VPS4 ATP hydrolysis nearly eliminated centriolar satellites and paused ciliogensis after formation of the ciliary vesicle. Zebrafish embryos injected with GFP-VPS4mRNA were less viable, exhibited developmental defects and had fewer cilia in Kupffer's vesicle. Surprisingly, ESCRT III proteins seldom localized to centrosomes and their depletion did not lead to these phenotypes. Our data support an ESCRT III-independent function for VPS4 at the centrosome and reveal that this evolutionary conserved AAA ATPase influences diverse centrosome functions and, as a result, global cellular architecture and development.
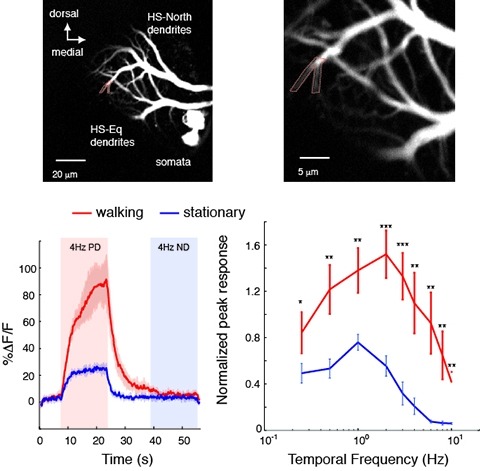
Changes in behavioral state modify neural activity in many systems. In some vertebrates such modulation has been observed and interpreted in the context of attention and sensorimotor coordinate transformations. Here we report state-dependent activity modulations during walking in a visual-motor pathway of Drosophila. We used two-photon imaging to monitor intracellular calcium activity in motion-sensitive lobula plate tangential cells (LPTCs) in head-fixed Drosophila walking on an air-supported ball. Cells of the horizontal system (HS)–a subgroup of LPTCs–showed stronger calcium transients in response to visual motion when flies were walking rather than resting. The amplified responses were also correlated with walking speed. Moreover, HS neurons showed a relatively higher gain in response strength at higher temporal frequencies, and their optimum temporal frequency was shifted toward higher motion speeds. Walking-dependent modulation of HS neurons in the Drosophila visual system may constitute a mechanism to facilitate processing of higher image speeds in behavioral contexts where these speeds of visual motion are relevant for course stabilization.
Nervous systems have evolved to translate external stimuli into appropriate behavioral responses. In an ever-changing environment, flexible adjustment of behavioral choice by experience-dependent learning is essential for the animal's survival. Associative learning is a simple form of learning that is widely observed from worms to humans. To understand the whole process of learning, we need to know how sensory information is represented and transformed in the brain, how it is changed by experience, and how the changes are reflected on motor output. To tackle these questions, studying numerically simple invertebrate nervous systems has a great advantage. In this review, I will feature the Pavlovian olfactory learning in the fruit fly, Drosophila melanogaster. The mushroom body is a key brain area for the olfactory learning in this organism. Recently, comprehensive anatomical information and the genetic tool sets were made available for the mushroom body circuit. This greatly accelerated the physiological understanding of the learning process. One of the key findings was dopamine-induced long-term synaptic plasticity that can alter the representations of stimulus valence. I will mostly focus on the new studies within these few years and discuss what we can possibly learn about the vertebrate systems from this model organism.
As most of us are aware, today's primary school, high school and undergraduate biology programs are struggling to incorporate even a fraction of the 'molecular revolution'of biological knowledge and technologies that surround us. In the first term alone, life science and biology classes of the new millennia routinely cover condensed versions of the year-long classes taught in the 60s, 70s and 80s. Teachers no longer have the luxury of spending half a year presenting Mendel and his peas.
Much work has explored animal-to-animal variability and compensation in ion channel expression. Yet, little is known regarding the physiological consequences of morphological variability. We quantify animal-to-animal variability in cable lengths (CV = 0.4) and branching patterns in the Gastric Mill (GM) neuron, an identified neuron type with highly-conserved physiological properties in the crustacean stomatogastric ganglion (STG) of \textitCancer borealis. We examined passive GM electrotonic structure by measuring the amplitudes and apparent reversal potentials (E\textsubscriptrevs) of inhibitory responses evoked with focal glutamate photo-uncaging in the presence of TTX. Apparent E\textsubscriptrevs were relatively invariant across sites (mean CV ± SD = 0.04 ± 0.01; 7–20 sites in each of 10 neurons), which ranged between 100–800 µm from the somatic recording site. Thus, GM neurons are remarkably electrotonically compact (estimated λ > 1.5 mm). Electrotonically compact structures, in consort with graded transmission, provide an elegant solution to observed morphological variability in the STG.
In this issue of Neuron, Rinberg et al. show that mice use a speed-accuracy tradeoff in odor discrimination. Shorter sampling results in high performance for easy problems, and enforced longer sampling results in higher accuracy for difficult problems, but mice freely choose intermediate sampling durations and accuracy varies with difficulty. Reward value and task requirements may determine sampling time choice and performance levels.