Filter
Associated Lab
- Aguilera Castrejon Lab (19) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (73) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Beyene Lab (15) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (55) Apply Branson Lab filter
- Card Lab (43) Apply Card Lab filter
- Cardona Lab (64) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (16) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (3) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (56) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (23) Apply Espinosa Medina Lab filter
- Feliciano Lab (12) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (30) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (65) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (30) Apply Hermundstad Lab filter
- Hess Lab (80) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (48) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (6) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (76) Apply Keller Lab filter
- Koay Lab (19) Apply Koay Lab filter
- Lavis Lab (158) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (32) Apply Li Lab filter
- Lippincott-Schwartz Lab (180) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (8) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (65) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (8) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (52) Apply Pachitariu Lab filter
- Pastalkova Lab (19) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (55) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (51) Apply Romani Lab filter
- Rubin Lab (149) Apply Rubin Lab filter
- Saalfeld Lab (64) Apply Saalfeld Lab filter
- Satou Lab (18) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (70) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (23) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (98) Apply Spruston Lab filter
- Stern Lab (160) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (41) Apply Stringer Lab filter
- Svoboda Lab (136) Apply Svoboda Lab filter
- Tebo Lab (35) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (53) Apply Turaga Lab filter
- Turner Lab (38) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (4) Apply Voigts Lab filter
- Wang (Meng) Lab (29) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (9) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2026 (70) Apply 2026 filter
- 2025 (222) Apply 2025 filter
- 2024 (210) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (192) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4265 Publications
Showing 2721-2730 of 4265 resultsReconstructing a connectome from an EM dataset often requires a large effort of proofreading automatically generated segmentations. While many tools exist to enable tracing or proofreading, recent advances in EM imaging and segmentation quality suggest new strategies and pose unique challenges for tool design to accelerate proofreading. Namely, we now have access to very large multi-TB EM datasets where (1) many segments are largely correct, (2) segments can be very large (several GigaVoxels), and where (3) several proofreaders and scientists are expected to collaborate simultaneously. In this paper, we introduce NeuTu as a solution to efficiently proofread large, high-quality segmentation in a collaborative setting. NeuTu is a client program of our high-performance, scalable image database called DVID so that it can easily be scaled up. Besides common features of typical proofreading software, NeuTu tames unprecedentedly large data with its distinguishing functions, including: (1) low-latency 3D visualization of large mutable segmentations; (2) interactive splitting of very large false merges with highly optimized semi-automatic segmentation; (3) intuitive user operations for investigating or marking interesting points in 3D visualization; (4) visualizing proofreading history of a segmentation; and (5) real-time collaborative proofreading with lock-based concurrency control. These unique features have allowed us to manage the workflow of proofreading a large dataset smoothly without dividing them into subsets as in other segmentation-based tools. Most importantly, NeuTu has enabled some of the largest connectome reconstructions as well as interesting discoveries in the fly brain.
Real-time lineage tracing in flies gets a boost with three techniques to specifically label a progenitor’s daughter cells.
How memories of past events influence behavior is a key question in neuroscience. The major associative learning center in Drosophila, the Mushroom Body (MB), communicates to the rest of the brain through Mushroom Body Output Neurons (MBONs). While 21 MBON cell types have their dendrites confined to small compartments of the MB lobes, analysis of EM connectomes revealed the presence of an additional 14 MBON cell types that are atypical in having dendritic input both within the MB lobes and in adjacent brain regions. Genetic reagents for manipulating atypical MBONs and experimental data on their functions has been lacking. In this report we describe new cell-type-specific GAL4 drivers for many MBONs, including the majority of atypical MBONs. Using these genetic reagents, we conducted optogenetic activation screening to examine their ability to drive behaviors and learning. These reagents provide important new tools for the study of complex behaviors in Drosophila.
Temporal focusing (TF) multiphoton systems constitute a powerful solution for cellular resolution optogenetic stimulation and recording in three-dimensional, scattering tissue. Here, we address two fundamental aspects in the design of such systems: first, we examine the design of TF systems with specific optical sectioning by comparatively analyzing previously published results. Next, we develop a solution for obtaining TF in a flexible three-dimensional pattern of cellmatched focal spots. Our solution employs spatio-temporal focusing (SSTF) in a unique optical system design that can be integrated before essentially any multiphoton imaging or stimulation system.
Chemical signaling between neurons in the brain can be divided into two major categories: fast synaptic transmission and neuromodulation. Fast synaptic transmission, mediated by amino acids such as glutamate and GABA, occurs on millisecond time scales and results in the influx of ions through ligand-gated ion channels on postsynaptic neurons (Figure 1A). Electrophysiological and optical imaging tools, including genetically encoded voltage indicators, have enabled neuroscientists to link cause (neurotransmitter release) and effect (membrane polarization) of synaptic transmission in time and space. Unlike classical neurotransmitters, neuromodulators do not produce immediate electrical effects that excite or inhibit target neurons. Instead, neuromodulators tune the intrinsic or synaptic properties of neurons, most commonly through interaction with G-protein-coupled receptors (GPCRs) (Figure 1B). Neuromodulators can escape the synaptic cleft and diffuse broadly, allowing them to influence the activity of many neurons in a state-dependent manner. Therefore, the spatial component of neuromodulator flux is fundamentally important. However, the temporal and/or spatial limitations of techniques classically used to study neuromodulation, such as microdialysis and fast-scan cyclic voltammetry (FSCV), make it difficult to interpret how neuromodulator release affects the plasticity or function of target neuronal populations on a moment-to-moment basis. Therefore, tools that can detect neuromodulators with high spatiotemporal resolution are critical for understanding their impact on neural computations that control behavior in health and disease.
2D template matching (2DTM) can be used to detect molecules and their assemblies in cellular cryo-EM images with high positional and orientational accuracy. While 2DTM successfully detects spherical targets such as large ribosomal subunits, challenges remain in detecting smaller and more aspherical targets in various environments. In this work, a novel 2DTM metric, referred to as the 2DTM p-value, is developed to extend the 2DTM framework to more complex applications. The 2DTM p-value combines information from two previously used 2DTM metrics, namely the 2DTM signal-to-noise ratio (SNR) and z-score, which are derived from the cross-correlation coefficient between the target and the template. The 2DTM p-value demonstrates robust detection accuracies under various imaging and sample conditions and outperforms the 2DTM SNR and z-score alone. Specifically, the 2DTM p-value improves the detection of aspherical targets such as a modified artificial tubulin patch particle (500 kDa) and a much smaller clathrin monomer (193 kDa) in simulated data. It also accurately recovers mature 60S ribosomes in yeast lamellae samples, even under conditions of increased Gaussian noise. The new metric will enable the detection of a wider variety of targets in both purified and cellular samples through 2DTM.
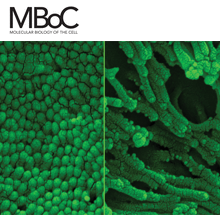
Visualization of cellular and molecular processes is an indispensable tool for cell biologists, and innovations in microscopy methods unfailingly lead to new biological discoveries. Today, light microscopy (LM) provides ever-higher spatial and temporal resolution and visualization of biological process over enormous ranges. Electron microscopy (EM) is moving into the atomic resolution regime and allowing cellular analyses that are more physiological and sophisticated in scope. Importantly, much is being gained by combining multiple approaches, (e.g., LM and EM) to take advantage of their complementary strengths. The advent of high-throughput microscopies has led to a common need for sophisticated computational methods to quantitatively analyze huge amounts of data and translate images into new biological insights.
Because of its genetic, molecular, and behavioral tractability, Drosophila has emerged as a powerful model system for studying molecular and cellular mechanisms underlying the development and function of nervous systems. The Drosophila nervous system has fewer neurons and exhibits a lower glia:neuron ratio than is seen in vertebrate nervous systems. Despite the simplicity of the Drosophila nervous system, glial organization in flies is as sophisticated as it is in vertebrates. Furthermore, fly glial cells play vital roles in neural development and behavior. In addition, powerful genetic tools are continuously being created to explore cell function in vivo. In taking advantage of these features, the fly nervous system serves as an excellent model system to study general aspects of glial cell development and function in vivo. In this article, we review and discuss advanced genetic tools that are potentially useful for understanding glial cell biology in Drosophila.
Multilocus sequence typing (MLST) has become the preferred method for genotyping many biological species, and it is especially useful for analyzing haploid eukaryotes. MLST is rigorous, reproducible, and informative, and MLST genotyping has been shown to identify major phylogenetic clades, molecular groups, or subpopulations of a species, as well as individual strains or clones. MLST molecular types often correlate with important phenotypes. Conventional MLST involves the extraction of genomic DNA and the amplification by PCR of several conserved, unlinked gene sequences from a sample of isolates of the taxon under investigation. In some cases, as few as three loci are sufficient to yield definitive results. The amplicons are sequenced, aligned, and compared by phylogenetic methods to distinguish statistically significant differences among individuals and clades. Although MLST is simpler, faster, and less expensive than whole genome sequencing, it is more costly and time-consuming than less reliable genotyping methods (e.g. amplified fragment length polymorphisms). Here, we describe a new MLST method that uses next-generation sequencing, a multiplexing protocol, and appropriate analytical software to provide accurate, rapid, and economical MLST genotyping of 96 or more isolates in single assay. We demonstrate this methodology by genotyping isolates of the well-characterized, human pathogenic yeast Cryptococcus neoformans.
NF-κB signaling has been implicated in neurodegenerative disease, epilepsy, and neuronal plasticity. However, the cellular and molecular activity of NF-κB signaling within the nervous system remains to be clearly defined. Here, we show that the NF-κB and IκB homologs Dorsal and Cactus surround postsynaptic glutamate receptor (GluR) clusters at the Drosophila NMJ. We then show that mutations in dorsal, cactus, and IRAK/pelle kinase specifically impair GluR levels, assayed immunohistochemically and electrophysiologically, without affecting NMJ growth, the size of the postsynaptic density, or homeostatic plasticity. Additional genetic experiments support the conclusion that cactus functions in concert with, rather than in opposition to, dorsal and pelle in this process. Finally, we provide evidence that Dorsal and Cactus act posttranscriptionally, outside the nucleus, to control GluR density. Based upon our data we speculate that Dorsal, Cactus, and Pelle could function together, locally at the postsynaptic density, to specify GluR levels.