Filter
Associated Lab
- Aguilera Castrejon Lab (19) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (75) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Beyene Lab (15) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (56) Apply Branson Lab filter
- Card Lab (43) Apply Card Lab filter
- Cardona Lab (64) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (16) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (3) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (58) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (25) Apply Espinosa Medina Lab filter
- Feliciano Lab (16) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (30) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (65) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (32) Apply Hermundstad Lab filter
- Hess Lab (80) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (49) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (7) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (15) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (77) Apply Keller Lab filter
- Koay Lab (20) Apply Koay Lab filter
- Lavis Lab (162) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (32) Apply Li Lab filter
- Lippincott-Schwartz Lab (182) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (9) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (65) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (8) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (56) Apply Pachitariu Lab filter
- Pastalkova Lab (19) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (55) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (52) Apply Romani Lab filter
- Rubin Lab (149) Apply Rubin Lab filter
- Saalfeld Lab (66) Apply Saalfeld Lab filter
- Satou Lab (18) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (72) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (23) Apply Sgro Lab filter
- Shroff Lab (35) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (98) Apply Spruston Lab filter
- Stern Lab (160) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (44) Apply Stringer Lab filter
- Svoboda Lab (136) Apply Svoboda Lab filter
- Tebo Lab (36) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (53) Apply Turaga Lab filter
- Turner Lab (38) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (5) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (11) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2026 (118) Apply 2026 filter
- 2025 (223) Apply 2025 filter
- 2024 (209) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (192) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4313 Publications
Showing 511-520 of 4313 resultsDescending neurons (DNs) occupy a key position in the sensorimotor hierarchy, conveying signals from the brain to the rest of the body below the neck. In Drosophila melanogaster flies, approximately 480 DN cell types have been described from electron-microscopy image datasets. Genetic access to these cell types is crucial for further investigation of their role in generating behaviour. We previously conducted the first large-scale survey of Drosophila melanogaster DNs, describing 98 unique cell types from light microscopy and generating cell-type-specific split-Gal4 driver lines for 65 of them. Here, we extend our previous work, describing the morphology of 137 additional DN types from light microscopy, bringing the total number DN types identified in light microscopy datasets to 235, or nearly 50%. In addition, we produced 500 new sparse split-Gal4 driver lines and compiled a list of previously published DN lines from the literature for a combined list of 738 split-Gal4 driver lines targeting 171 DN types.
Simultaneous recordings from hundreds or thousands of neurons are becoming routine because of innovations in instrumentation, molecular tools, and data processing software. Such recordings can be analyzed with data science methods, but it is not immediately clear what methods to use or how to adapt them for neuroscience applications. We review, categorize, and illustrate diverse analysis methods for neural population recordings and describe how these methods have been used to make progress on longstanding questions in neuroscience. We review a variety of approaches, ranging from the mathematically simple to the complex, from exploratory to hypothesis-driven, and from recently developed to more established methods. We also illustrate some of the common statistical pitfalls in analyzing large-scale neural data.
Flexible goal-driven orientation requires that the position of a target be stored, especially in case the target moves out of sight. The capability to retain, recall and integrate such positional information into guiding behaviour has been summarized under the term spatial working memory. This kind of memory contains specific details of the presence that are not necessarily part of a long-term memory. Neurophysiological studies in primates indicate that sustained activity of neurons encodes the sensory information even though the object is no longer present. Furthermore they suggest that dopamine transmits the respective input to the prefrontal cortex, and simultaneous suppression by GABA spatially restricts this neuronal activity. Here we show that Drosophila melanogaster possesses a similar spatial memory during locomotion. Using a new detour setup, we show that flies can remember the position of an object for several seconds after it has been removed from their environment. In this setup, flies are temporarily lured away from the direction towards their hidden target, yet they are thereafter able to aim for their former target. Furthermore, we find that the GABAergic (stainable with antibodies against GABA) ring neurons of the ellipsoid body in the central brain are necessary and their plasticity is sufficient for a functional spatial orientation memory in flies. We also find that the protein kinase S6KII (ignorant) is required in a distinct subset of ring neurons to display this memory. Conditional expression of S6KII in these neurons only in adults can restore the loss of the orientation memory of the ignorant mutant. The S6KII signalling pathway therefore seems to be acutely required in the ring neurons for spatial orientation memory in flies.
The C. elegans cell lineage provides a unique opportunity to look at how cell lineage affects patterns of gene expression. We developed an automatic cell lineage analyzer that converts high-resolution images of worms into a data table showing fluorescence expression with single-cell resolution. We generated expression profiles of 93 genes in 363 specific cells from L1 stage larvae and found that cells with identical fates can be formed by different gene regulatory pathways. Molecular signatures identified repeating cell fate modules within the cell lineage and enabled the generation of a molecular differentiation map that reveals points in the cell lineage when developmental fates of daughter cells begin to diverge. These results demonstrate insights that become possible using computational approaches to analyze quantitative expression from many genes in parallel using a digital gene expression atlas.

The rhodopsin genes of Drosophila melanogaster are expressed in nonoverlapping subsets of photoreceptor cells within the insect visual system. Two of these genes, Rh3 and Rh4, are known to display complementary expression patterns in the UV-sensitive R7 photoreceptor cell population of the compound eye. In addition, we find that Rh3 is expressed in a small group of paired R7 and R8 photoreceptor cells at the dorsal eye margin that are apparently specialized for the detection of polarized light. In this paper we present a detailed characterization of the cis-acting requirements of both Rh3 and Rh4. Promoter deletion series demonstrate that small regulatory regions (less than 300 bp) of both R7 opsin genes contain DNA sequences sufficient to generate their respective expression patterns. Individual cis-acting elements were further identified by oligonucleotide-directed mutagenesis guided by interspecific sequence comparisons. Our results suggest that the Drosophila rhodopsin genes share a simple bipartite promoter structure, whereby the proximal region constitutes a functionally equivalent promoter "core" and the distal region determines cell-type specificity. The expression patterns of several hybrid rhodopsin promoters, in which all or part of the putative core regions have been replaced with the analogous regions of different rhodopsin promoters, provide additional evidence in support of this model.
e13028Background: Liquid biopsy has emerged as a powerful, minimally invasive tool for predicting treatment response and survival in breast and other advanced cancers. However, the detection and characterization of circulating tumor cells (CTCs) — a key factor in metastatic progression—remain challenging due to their low frequency and reliance on manual, time-intensive validation using only a couple of established methods for immunofluorescence staining, such as CellSearch. Harnessing deep learning for automated CTC detection and characterization of the blood cells interacting with CTCs holds the potential to advance prognostic evaluations and guide more effective therapies significantly. Methods: Leveraging FDA-approved CellSearch technology and sequencing approaches, we analyzed 2,853 blood specimens, longitudinally collected from 1358 patients with advanced cancer (breast, prostate, etc) and additional diseases. We built a novel deep learning platform, CTCpose, which integrates machine learning and AI-driven image analysis to automate the detection and categorization of CTCs, white blood cells (WBCs), and their clustering interactions. We extracted cellular and nuclear features to enable precise evaluation of individual CTCs, WBCs, homotypic CTC clusters, heterotypic CTC–WBC clusters, and immune cell aggregates. Results: By employing the CTCpose platform, we achieved fully automated identification of CTCs and immune cells, unraveling the spatial organization and functional characteristics of both homotypic and heterotypic clusters. These highly granular assessments revealed clinically significant correlations with patient survival, disease progression, and therapeutic outcomes. Our data underscore the critical role of CTC–immune cell interactions and the dynamic shifts in CTC phenotypes—both as single cells and clusters—in stratifying patients by risk and informing treatment strategies. Conclusions: This work illustrates the transformative power of deep learning in the analysis of liquid biopsy samples. By overcoming the limitations of traditional CTC detection, we have established a robust framework that integrates imaging data with large-scale patient cohorts to deliver predictive models of high clinical relevance. The CTCpose platform not only refines our understanding of CTC–immune cell biology but also paves the way for personalized oncology approaches, highlighting the impactful convergence of artificial intelligence and precision medicine.
Regulation of transcription during embryogenesis is key to development and differentiation. To study transcript expression throughout Caenorhabditis elegans embryogenesis at single-molecule resolution, we developed a high-throughput single-molecule fluorescence in situ hybridization (smFISH) method that relies on computational methods to developmentally stage embryos and quantify individual mRNA molecules in single embryos. We applied our system to sdc-2, a zygotically transcribed gene essential for hermaphrodite development and dosage compensation. We found that sdc-2 is rapidly activated during early embryogenesis by increasing both the number of mRNAs produced per transcription site and the frequency of sites engaged in transcription. Knockdown of sdc-2 and dpy-27, a subunit of the dosage compensation complex (DCC), increased the number of active transcription sites for the X chromosomal gene dpy-23 but not the autosomal gene mdh-1, suggesting that the DCC reduces the frequency of dpy-23 transcription. The temporal resolution from in silico staging of embryos showed that the deletion of a single DCC recruitment element near the dpy-23 gene causes higher dpy-23 mRNA expression after the start of dosage compensation, which could not be resolved using mRNAseq from mixed-stage embryos. In summary, we have established a computational approach to quantify temporal regulation of transcription throughout C. elegans embryogenesis and demonstrated its potential to provide new insights into developmental gene regulation.
Regulation of transcription during embryogenesis is key to development and differentiation. To study transcript expression throughout Caenorhabditis elegans embryogenesis at single-molecule resolution, we developed a high-throughput single-molecule fluorescence in situ hybridization (smFISH) method that relies on computational methods to developmentally stage embryos and quantify individual mRNA molecules in single embryos. We applied our system to sdc-2, a zygotically transcribed gene essential for hermaphrodite development and dosage compensation. We found that sdc-2 is rapidly activated during early embryogenesis by increasing both the number of mRNAs produced per transcription site and the frequency of sites engaged in transcription. Knockdown of sdc-2 and dpy-27, a subunit of the dosage compensation complex (DCC), increased the number of active transcription sites for the X chromosomal gene dpy-23 but not the autosomal gene mdh-1, suggesting that the DCC reduces the frequency of dpy-23 transcription. The temporal resolution from in silico staging of embryos showed that the deletion of a single DCC recruitment element near the dpy-23 gene causes higher dpy-23 mRNA expression after the start of dosage compensation, which could not be resolved using mRNAseq from mixed-stage embryos. In summary, we have established a computational approach to quantify temporal regulation of transcription throughout C. elegans embryogenesis and demonstrated its potential to provide new insights into developmental gene regulation.Competing Interest StatementThe authors have declared no competing interest.
Single-particle electron cryo-microscopy and computational image classification can be used to analyze structural variability in macromolecules and their assemblies. In some cases, a particle may contain different regions that each display a range of distinct conformations. We have developed strategies, implemented within the Frealign and cisTEM image processing packages, to focus classify on specific regions of a particle and detect potential covariance. The strategies are based on masking the region of interest using either a 2-D mask applied to reference projections and particle images, or a 3-D mask applied to the 3-D volume. We show that focused classification approaches can be used to study structural covariance, a concept that is likely to gain more importance as datasets grow in size, allowing the distinction of more structural states and smaller differences between states. Finally, we apply the approaches to an experimental dataset containing the HIV-1 Transactivation Response (TAR) element RNA fused into the large bacterial ribosomal subunit to deconvolve structural mobility within localized regions of interest, and to a dataset containing assembly intermediates of the large subunit to measure structural covariance.
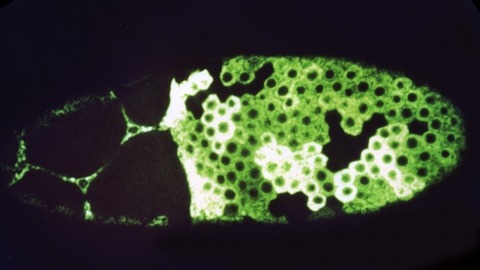
We have constructed a series of strains to facilitate the generation and analysis of clones of genetically distinct cells in developing and adult tissues of Drosophila. Each of these strains carries an FRT element, the target for the yeast FLP recombinase, near the base of a major chromosome arm, as well as a gratuitous cell-autonomous marker. Novel markers that carry epitope tags and that are localized to either the cell nucleus or cell membrane have been generated. As a demonstration of how these strains can be used to study a particular gene, we have analyzed the developmental role of the Drosophila EGF receptor homolog. Moreover, we have shown that these strains can be utilized to identify new mutations in mosaic animals in an efficient and unbiased way, thereby providing an unprecedented opportunity to perform systematic genetic screens for mutations affecting many biological processes.