Filter
Associated Lab
- Aguilera Castrejon Lab (4) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (64) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (104) Apply Betzig Lab filter
- Beyene Lab (10) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (53) Apply Branson Lab filter
- Card Lab (37) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (15) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dennis Lab (2) Apply Dennis Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (46) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (21) Apply Espinosa Medina Lab filter
- Feliciano Lab (16) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (17) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (55) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (28) Apply Hermundstad Lab filter
- Hess Lab (77) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (45) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (2) Apply Johnson Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (62) Apply Keller Lab filter
- Koay Lab (4) Apply Koay Lab filter
- Lavis Lab (150) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (8) Apply Li Lab filter
- Lippincott-Schwartz Lab (110) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (4) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (60) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (7) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (44) Apply Pachitariu Lab filter
- Pastalkova Lab (6) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (49) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (40) Apply Romani Lab filter
- Rubin Lab (111) Apply Rubin Lab filter
- Saalfeld Lab (49) Apply Saalfeld Lab filter
- Satou Lab (3) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (55) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (3) Apply Sgro Lab filter
- Shroff Lab (35) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (62) Apply Spruston Lab filter
- Stern Lab (77) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (41) Apply Stringer Lab filter
- Svoboda Lab (132) Apply Svoboda Lab filter
- Tebo Lab (12) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (19) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (41) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (5) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (11) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (49) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (18) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- High Performance Computing (7) Apply High Performance Computing filter
- Integrative Imaging (25) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (40) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Mass Spectrometry (1) Apply Mass Spectrometry filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Project Technical Resources (54) Apply Project Technical Resources filter
- Quantitative Genomics (20) Apply Quantitative Genomics filter
- Scientific Computing (106) Apply Scientific Computing filter
- Stem Cell & Primary Culture (14) Apply Stem Cell & Primary Culture filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2026 (118) Apply 2026 filter
- 2025 (223) Apply 2025 filter
- 2024 (208) Apply 2024 filter
- 2023 (157) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2896 Janelia Publications
Showing 1511-1520 of 2896 resultsManifold attractors are a key framework for understanding how continuous variables, such as position or head direction, are encoded in the brain. In this framework, the variable is represented along a continuum of persistent neuronal states which forms a manifold attactor. Neural networks with symmetric synaptic connectivity that can implement manifold attractors have become the dominant model in this framework. In addition to a symmetric connectome, these networks imply homogeneity of individual-neuron tuning curves and symmetry of the representational space; these features are largely inconsistent with neurobiological data. Here, we developed a theory for computations based on manifold attractors in trained neural networks and show how these manifolds can cope with diverse neuronal responses, imperfections in the geometry of the manifold and a high level of synaptic heterogeneity. In such heterogeneous trained networks, a continuous representational space emerges from a small set of stimuli used for training. Furthermore, we find that the network response to external inputs depends on the geometry of the representation and on the level of synaptic heterogeneity in an analytically tractable and interpretable way. Finally, we show that a too complex geometry of the neuronal representation impairs the attractiveness of the manifold and may lead to its destabilization. Our framework reveals that continuous features can be represented in the recurrent dynamics of heterogeneous networks without assuming unrealistic symmetry. It suggests that the representational space of putative manifold attractors in the brain dictates the dynamics in their vicinity.
Cortical neurons form specific circuits, but the functional structure of this microarchitecture and its relation to behaviour are poorly understood. Two-photon calcium imaging can monitor activity of spatially defined neuronal ensembles in the mammalian cortex. Here we applied this technique to the motor cortex of mice performing a choice behaviour. Head-fixed mice were trained to lick in response to one of two odours, and to withhold licking for the other odour. Mice routinely showed significant learning within the first behavioural session and across sessions. Microstimulation and trans-synaptic tracing identified two non-overlapping candidate tongue motor cortical areas. Inactivating either area impaired voluntary licking. Imaging in layer 2/3 showed neurons with diverse response types in both areas. Activity in approximately half of the imaged neurons distinguished trial types associated with different actions. Many neurons showed modulation coinciding with or preceding the action, consistent with their involvement in motor control. Neurons with different response types were spatially intermingled. Nearby neurons (within approximately 150 mum) showed pronounced coincident activity. These temporal correlations increased with learning within and across behavioural sessions, specifically for neuron pairs with similar response types. We propose that correlated activity in specific ensembles of functionally related neurons is a signature of learning-related circuit plasticity. Our findings reveal a fine-scale and dynamic organization of the frontal cortex that probably underlies flexible behaviour.
Two simple models—vaulting over stiff legs and rebounding over compliant legs—are employed to describe the mechanics of legged locomotion. It is agreed that compliant legs are necessary for describing running, and that leg compliance is also present during walking. Stiff legs continue to be employed to model walking under the assumption that the compliance of the leg during walking is low enough to be considered stiff. Here we study gait choice and walk-to-run transition in a biped with compliance and show that the principles underlying gait choice and transition are completely different from stiff legs. Two findings underpin our conclusions: First, at the same speed, step length and stance duration, multiple gaits that differ in the number of times the leg expands and contracts during a single stance are possible. Among them, humans and other animals choose the (normal) gait with M-shaped vertical ground reaction forces (vGRF) not just because of energy considerations but also constraints from forces. Second, the transition from walking to running occurs because of three factors: vGRF minimum at mid-stance characteristic of normal walking, synchronization of horizontal and vertical motions during single support, and velocity redirection during the double support. The insight above required an analytical approximation of the double spring-loaded pendulum (DSLIP) model describing the intricate oscillatory dynamics that relate single and double support phases. Additionally, we also examined DSLIP as a quantitative model for locomotion and conclude that DSLIP speed range is limited. However, insights gleaned from the analytical treatment of DSLIP are general and will inform the construction of more accurate models of walking. bioRxiv preprint: https://doi.org/10.1101/2024.09.23.612940
Leptin is an adipose tissue hormone that maintains homeostatic control of adipose tissue mass by regulating the activity of specific neural populations controlling appetite and metabolism1. Leptin regulates food intake by inhibiting orexigenic agouti-related protein (AGRP) neurons and activating anorexigenic pro-opiomelanocortin (POMC) neurons2. However, while AGRP neurons regulate food intake on a rapid time scale, acute activation of POMC neurons has only a minimal effect3–5. This has raised the possibility that there is a heretofore unidentified leptin-regulated neural population that suppresses appetite on a rapid time scale. Here, we report the discovery of a novel population of leptin-target neurons expressing basonuclin 2 (Bnc2) that acutely suppress appetite by directly inhibiting AGRP neurons. Opposite to the effect of AGRP activation, BNC2 neuronal activation elicited a place preference indicative of positive valence in hungry but not fed mice. The activity of BNC2 neurons is finely tuned by leptin, sensory food cues, and nutritional status. Finally, deleting leptin receptors in BNC2 neurons caused marked hyperphagia and obesity, similar to that observed in a leptin receptor knockout in AGRP neurons. These data indicate that BNC2-expressing neurons are a key component of the neural circuit that maintains energy balance, thus filling an important gap in our understanding of the regulation of food intake and leptin action.
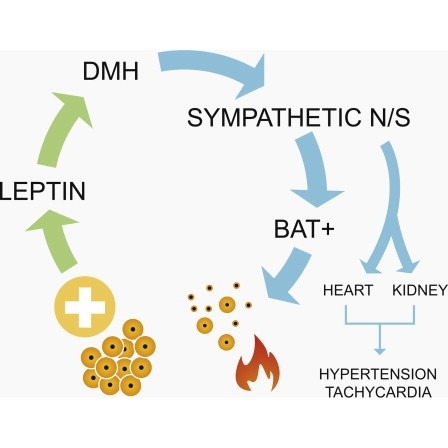
Obesity is associated with increased blood pressure (BP), which in turn increases the risk of cardiovascular diseases. We found that the increase in leptin levels seen in diet-induced obesity (DIO) drives an increase in BP in rodents, an effect that was not seen in animals deficient in leptin or leptin receptors (LepR). Furthermore, humans with loss-of-function mutations in leptin and the LepR have low BP despite severe obesity. Leptin's effects on BP are mediated by neuronal circuits in the dorsomedial hypothalamus (DMH), as blocking leptin with a specific antibody, antagonist, or inhibition of the activity of LepR-expressing neurons in the DMH caused a rapid reduction of BP in DIO mice, independent of changes in weight. Re-expression of LepRs in the DMH of DIO LepR-deficient mice caused an increase in BP. These studies demonstrate that leptin couples changes in weight to changes in BP in mammalian species.
Although the vinegar fly, Drosophila melanogaster, has been a biological model organism for over a century, its emergence as a model system for the study of neurophysiology is comparatively recent. The primary reason for this is that the vinegar fly and its neurons are tiny; up until 5 years ago, it was prohibitively difficult to record intracellularly from individual neurons in the intact Drosophila brain (Wilson et al., 2004). Today, fly electrophysiologists can genetically label neurons with GFP and reliably record from many (but not all) neurons in the fruit fly brain. Using genetic tools to drive expression of fluorescent calcium indicators, light-sensitive ion channels, or cell activity suppressors, we are beginning to understand how the external environment is represented with electrical potentials in Drosophila neurons (for review, see Olsen and Wilson, 2008).
Natural neural circuits, optimized by millions of years of evolution, are fast, low power, robust, and adapt in response to experience, all characteristics we would love to have in systems we ourselves design. Recently there have been enormous advances in understanding how neurons implement computations within the brain of living creatures. Can we use this new-found knowledge to create better artificial system? What lessons can we learn from the neurons themselves, that can help us create better neuromorphic circuits?
Upon inflammation, leukocytes extravasate through endothelial cells. When they extravasate, it is generally accepted that neighboring endothelial cells disconnect. Careful examination of endothelial junctions showed a partial membrane overlap beyond VE-cadherin distribution. These overlaps are regulated by actin polymerization and, although marked by, do not require PECAM-1, nor VE-cadherin. Neutrophils prefer wider membrane overlaps as exit sites. Detailed 3D analysis of neutrophil transmigration in real time at high spatiotemporal resolution revealed that overlapping endothelial membranes form a tunnel during neutrophil transmigration. These tunnels are formed by the neutrophil lifting the membrane of the upper endothelial cell while indenting and crawling over the membrane of the underlying endothelial cell. Our work shows that endothelial cells do not simply retract upon the passage of neutrophils but provide membrane tunnels, allowing neutrophils to extravasate. This discovery defines the 3D multicellular architecture in which the paracellular transmigration of neutrophils occurs.
Light sheet fluorescence microscopy (LSFM) uses a thin sheet of light to excite only fluorophores within the focal volume. Light sheet microscopes (LSMs) have a true optical sectioning capability and, hence, provide axial resolution, restrict photobleaching and phototoxicity to a fraction of the sample and use cameras to record tens to thousands of images per second. LSMs are used for in-depth analyses of large, optically cleared samples and long-term three-dimensional (3D) observations of live biological specimens at high spatio-temporal resolution. The independently operated illumination and detection trains and the canonical implementations, selective/single plane illumination microscope (SPIM) and digital scanned laser microscope (DSLM), are the basis for many LSM designs. In this Primer, we discuss various applications of LSFM for imaging multicellular specimens, developing vertebrate and invertebrate embryos, brain and heart function, 3D cell culture models, single cells, tissue sections, plants, organismic interaction and entire cleared brains. Further, we describe the combination of LSFM with other imaging approaches to allow for super-resolution or increased penetration depth and the use of sophisticated spatio-temporal manipulations to allow for observations along multiple directions. Finally, we anticipate developments of the field in the near future.
Capturing dynamic processes in live samples is a nontrivial task in biological imaging. Although fluorescence provides high specificity and contrast compared to other light microscopy techniques, the photophysical principles of this method can have a harmful effect on the sample. Current advances in light sheet microscopy have created a novel imaging toolbox that allows for rapid acquisition of high-resolution fluorescent images with minimal perturbation of the processes of interest. Each unique design has its own advantages and limitations. In this review, we describe several cutting edge light sheet microscopes and their optimal applications.