Filter
Associated Lab
- Aguilera Castrejon Lab (2) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (58) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (103) Apply Betzig Lab filter
- Beyene Lab (10) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (51) Apply Branson Lab filter
- Card Lab (37) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (41) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (18) Apply Espinosa Medina Lab filter
- Feliciano Lab (10) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (42) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (55) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (26) Apply Hermundstad Lab filter
- Hess Lab (76) Apply Hess Lab filter
- Ilanges Lab (3) Apply Ilanges Lab filter
- Jayaraman Lab (44) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (3) Apply Koay Lab filter
- Lavis Lab (144) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (6) Apply Li Lab filter
- Lippincott-Schwartz Lab (107) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (2) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (59) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (39) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (49) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (39) Apply Romani Lab filter
- Rubin Lab (110) Apply Rubin Lab filter
- Saalfeld Lab (47) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (52) Apply Schreiter Lab filter
- Sgro Lab (2) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (61) Apply Spruston Lab filter
- Stern Lab (75) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (36) Apply Stringer Lab filter
- Svoboda Lab (132) Apply Svoboda Lab filter
- Tebo Lab (11) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (18) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (41) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (4) Apply Voigts Lab filter
- Wang (Meng) Lab (27) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (9) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (41) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (18) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- High Performance Computing (7) Apply High Performance Computing filter
- Integrative Imaging (18) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (37) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Mass Spectrometry (1) Apply Mass Spectrometry filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (14) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (53) Apply Project Technical Resources filter
- Quantitative Genomics (20) Apply Quantitative Genomics filter
- Scientific Computing (100) Apply Scientific Computing filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2026 (17) Apply 2026 filter
- 2025 (225) Apply 2025 filter
- 2024 (211) Apply 2024 filter
- 2023 (157) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2800 Janelia Publications
Showing 1931-1940 of 2800 resultsA parallel wavefront optimization method is demonstrated experimentally to focus light through random scattering media. The simultaneous modulation of multiple phase elements, each at a unique frequency, enables a parallel determination of the optimal wavefront. Compared to a pixel-by-pixel measurement, the reported parallel method uses the target signal in a highly efficient way. With 441 phase elements, a high-quality focus was formed through a glass diffuser with a peak-to-background ratio of \~{}270. The accuracy and repeatability of the system were tested through experiments.
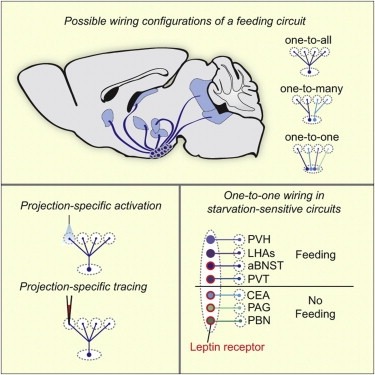
Neural circuits for essential natural behaviors are shaped by selective pressure to coordinate reliable execution of flexible goal-directed actions. However, the structural and functional organization of survival-oriented circuits is poorly understood due to exceptionally complex neuroanatomy. This is exemplified by AGRP neurons, which are a molecularly defined population that is sufficient to rapidly coordinate voracious food seeking and consumption behaviors. Here, we use cell-type-specific techniques for neural circuit manipulation and projection-specific anatomical analysis to examine the organization of this critical homeostatic circuit that regulates feeding. We show that AGRP neuronal circuits use a segregated, parallel, and redundant output configuration. AGRP neuron axon projections that target different brain regions originate from distinct subpopulations, several of which are sufficient to independently evoke feeding. The concerted anatomical and functional analysis of AGRP neuron projection populations reveals a constellation of core forebrain nodes, which are part of an extended circuit that mediates feeding behavior.
Wistar-Kyoto (WKY) rats as an endogenous depression model partially lack a response to classic selective serotonin reuptake inhibitors (SSRIs). Thus, this strain has the potential to be established as a model of treatment-resistant depression (TRD). However, the SSRI resistance in WKY rats is still not fully understood. In this study, WKY and control rats were subjected to a series of tests, namely, a forced swim test (FST), a sucrose preference test (SPT), and an open field test (OFT), and were scanned in a 7.0-T MRI scanner before and after three-week citalopram or saline administration. Behavioral results demonstrated that WKY rats had increased immobility in the FST and decreased sucrose preference in the SPT and central time spent in the OFT. However, citalopram did not improve immobility in the FST. The amplitude of low-frequency fluctuation (ALFF) analysis showed regional changes in the striatum and hippocampus of WKY rats. However, citalopram partially reversed the ALFF value in the dorsal part of the two regions. Functional connectivity (FC) analysis showed that FC strengths were decreased in WKY rats compared with controls. Nevertheless, citalopram partially increased FC strengths in WKY rats. Based on FC, global graph analysis demonstrated decreased network efficiency in WKY + saline group compared with control + saline group, but citalopram showed weak network efficiency improvement. In conclusion, resting-state fMRI results implied widely affected brain function at both regional and global levels in WKY rats. Citalopram had only partial effects on these functional changes, indicating a potential treatment resistance mechanism.
Significant technical challenges exist when measuring synaptic connections between neurons in living brain tissue. The patch clamping technique, when used to probe for synaptic connections, is manually laborious and time-consuming. To improve its efficiency, we pursued another approach: instead of retracting all patch clamping electrodes after each recording attempt, we cleaned just one of them and reused it to obtain another recording while maintaining the others. With one new patch clamp recording attempt, many new connections can be probed. By placing one pipette in front of the others in this way, one can 'walk' across the mouse brain slice, termed 'patch-walking.' We performed 136 patch clamp attempts for two pipettes, achieving 71 successful whole cell recordings (52.2%). Of these, we probed 29 pairs (i.e. 58 bidirectional probed connections) averaging 91 μm intersomatic distance, finding three connections. Patch-walking yields 80-92% more probed connections, for experiments with 10-100 cells than the traditional synaptic connection searching method.
Input comparison is thought to occur in many neuronal circuits, including the hippocampus, where functionally important interactions between the Schaffer collateral and perforant pathways have been hypothesized. We investigated this idea using multisite, whole-cell recordings and Ca2+ imaging and found that properly timed, repetitive stimulation of both pathways results in the generation of large plateau potentials in distal dendrites of CA1 pyramidal neurons. These dendritic plateau potentials produce widespread Ca2+ influx, large after-depolarizations, burst firing output, and long-term potentiation of perforant path synapses. Plateau duration is directly related to the strength and temporal overlap of pathway activation and involves back-propagating action potentials and both NMDA receptors and voltage-gated Ca2+ channels. Thus, the occurrence of highly correlated SC and PP input to CA1 is signaled by a dramatic change in output mode and an increase in input efficacy, all induced by a large plateau potential in the distal dendrites of CA1 pyramidal neurons.
Larval Drosophila offer a study case for behavioral neurogenetics that is simple enough to be experimentally tractable, yet complex enough to be worth the effort. We provide a detailed, hands-on manual for Pavlovian odor-reward learning in these animals. Given the versatility of Drosophila for genetic analyses, combined with the evolutionarily shared genetic heritage with humans, the paradigm has utility not only in behavioral neurogenetics and experimental psychology, but for translational biomedicine as well. Together with the upcoming total synaptic connectome of the Drosophila nervous system and the possibilities of single-cell-specific transgene expression, it offers enticing opportunities for research. Indeed, the paradigm has already been adopted by a number of labs and is robust enough to be used for teaching in classroom settings. This has given rise to a demand for a detailed, hands-on manual directed at newcomers and/or at laboratory novices, and this is what we here provide. The paradigm and the present manual have a unique set of features: • The paradigm is cheap, easy, and robust; • The manual is detailed enough for newcomers or laboratory novices; • It briefly covers the essential scientific context; • It includes sheets for scoring, data analysis, and display; • It is multilingual: in addition to an English version we provide German, French, Japanese, Spanish and Italian language versions as well. The present manual can thus foster science education at an earlier age and enable research by a broader community than has been the case to date.
The regular distribution of mitochondrial DNA-containing nucleoids is essential for mitochondrial function and genome inheritance; however, the underlying mechanisms remain unknown. Our data reveal that mitochondria frequently undergo spontaneous and reversible pearling - a biophysical instability in which tubules undulate into regularly spaced beads. We discovered that pearling imposes a characteristic length scale, simultaneously mediating nucleoid disaggregation and establishing inter-nucleoid distancing with near-maximally achievable precision. Cristae invaginations play a dual role: lamellar cristae density determines pearling frequency and duration, and preserves the resulting nucleoid spacing after recovery. The distribution of mitochondrial genomes is thus fundamentally governed by the interplay between spontaneous pearling and cristae ultrastructure.