Filter
Associated Lab
- Aguilera Castrejon Lab (4) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (64) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (104) Apply Betzig Lab filter
- Beyene Lab (10) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (53) Apply Branson Lab filter
- Card Lab (37) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (15) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dennis Lab (2) Apply Dennis Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (46) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (21) Apply Espinosa Medina Lab filter
- Feliciano Lab (16) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (17) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (55) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (28) Apply Hermundstad Lab filter
- Hess Lab (77) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (45) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (2) Apply Johnson Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (62) Apply Keller Lab filter
- Koay Lab (4) Apply Koay Lab filter
- Lavis Lab (150) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (8) Apply Li Lab filter
- Lippincott-Schwartz Lab (110) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (4) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (60) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (7) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (44) Apply Pachitariu Lab filter
- Pastalkova Lab (6) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (49) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (40) Apply Romani Lab filter
- Rubin Lab (111) Apply Rubin Lab filter
- Saalfeld Lab (49) Apply Saalfeld Lab filter
- Satou Lab (3) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (55) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (3) Apply Sgro Lab filter
- Shroff Lab (35) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (62) Apply Spruston Lab filter
- Stern Lab (77) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (41) Apply Stringer Lab filter
- Svoboda Lab (132) Apply Svoboda Lab filter
- Tebo Lab (12) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (19) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (41) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (5) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (11) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (49) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (18) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- High Performance Computing (7) Apply High Performance Computing filter
- Integrative Imaging (25) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (40) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Mass Spectrometry (1) Apply Mass Spectrometry filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Project Technical Resources (54) Apply Project Technical Resources filter
- Quantitative Genomics (20) Apply Quantitative Genomics filter
- Scientific Computing (106) Apply Scientific Computing filter
- Stem Cell & Primary Culture (14) Apply Stem Cell & Primary Culture filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2026 (118) Apply 2026 filter
- 2025 (223) Apply 2025 filter
- 2024 (208) Apply 2024 filter
- 2023 (157) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2896 Janelia Publications
Showing 1-10 of 2896 resultsSingle-stranded DNA (ssDNA)-functionalized single-wall carbon nanotubes (SWCNTs) exhibit exceptional optical sensitivity to catecholamines, including dopamine and norepinephrine—key signaling molecules that play vital roles in brain function. This unique capability positions SWCNTs as powerful tools for advancing our understanding of neurochemical processes involving dopaminergic and noradrenergic neurons. In this presentation, I will highlight how our lab has leveraged SWCNT nanosensors to push the boundaries of dopamine neuroscience. For studies in cultured neurons, we developed a composite nanofilm strategy that enabled us to visualize dopamine release with exceptional resolution, capturing single bouton activity with quantal sensitivity while monitoring thousands of release sites simultaneously in large imaging fields of view. By combining SWCNT-based activity imaging with immunofluorescence, electron microscopy, and cutting-edge molecular, cellular and genetic techniques, we have gained new insights into neurobiological properties of dopamine release sites in dopaminergic neurons that had heretofore been inaccessible with conventional methods of inquiry. Building on these advances, I will discuss recent progress in the development of in vivo-compatible dopamine nanosensors. These innovations have allowed us to monitor dopamine dynamics in awake and behaving mice, bridging the gap between molecular-scale imaging and real-time behavior analysis. Furthermore, I will discuss methodological developments that enabled the deployment of these nanosensors in vivo. Looking ahead, these SWCNT-enabled technological advancements hold potential for the study of neurochemical signaling, offering deeper insights into both normal brain function and the pathophysiology of disorders involving catecholamines. Future work aims to expand the applications of these nanosensors to other neural circuits and neuromodulators, ultimately advancing our ability to decode the brain’s chemical language.
Sleep is regulated by a homeostatic process and associated with an increased arousal threshold, but the genetic and neuronal mechanisms that implement these essential features of sleep remain poorly understood.To address these fundamental questions, we performed a zebrafish genetic screen informed by human genome-wide association studies.We found that mutation of serine/threonine kinase 32a (stk32a) results in increased sleep and impaired sleep homeostasis in both zebrafish and mice, and that stk32a acts downstream of neurotensin signaling and the serotonergic raphe in zebrafish. stk32a mutation reduces phosphorylation of neurofilament proteins, which are co-expressed with stk32a in neurons that regulate motor activity and in lateral line hair cells that detect environmental stimuli, and ablating these cells phenocopies stk32a mutation. Neurotensin signaling inhibits specific sensory and motor populations, and blocks stimulus-evoked responses of neurons that relay sensory information from hair cells to the brain.Our work thus shows that stk32a is an evolutionarily conserved sleep regulator that links neuropeptidergic and neuromodulatory systems to homeostatic sleep drive and changes in arousal threshold, which are implemented through suppression of specific sensory and motor systems.
Small-molecule fluorescent stains enable the imaging of cellular structures without the need for genetic manipulation. Here, we introduce 2,7-diaminobenzopyrylium (DAB) dyes as live-cell mitochondrial stains excited with violet light. This amalgam of the coumarin and rhodamine fluorophore structures yields dyes with high photostability and tunable spectral properties.
Mechanosensory corpuscles detect transient touch and vibratory signals in the skin of vertebrates, enabling navigation, foraging, and precise manipulation of objects1. The corpuscle core comprises a terminal neurite of a mechanoreceptor afferent, the only known touch-sensing element within corpuscles, surrounded by terminal Schwann cells called lamellar cells (LCs)2–4. However, the precise corpuscular ultrastructure, and the role of LCs in touch detection are unknown. Here we used enhanced focused ion beam scanning electron microscopy and electron tomography to reveal the three-dimensional architecture of avian Meissner (Grandry) corpuscle5. We show that corpuscles contain a stack of LCs innervated by two afferents, which form large-area contacts with LCs. LCs form tether-like connections with the afferent membrane and contain dense core vesicles which release their content onto the afferent. Furthermore, by performing simultaneous electrophysiological recordings from both cell types, we show that mechanosensitive LCs use calcium influx to trigger action potential firing in the afferent and thus serve as physiological touch sensors in the skin. Our findings suggest a bi-cellular mechanism of touch detection, which comprises the afferent and LCs, likely enables corpuscles to encode the nuances of tactile stimuli.
To image the accessible genome at nanometer scale in situ, we developed three-dimensional assay for transposase-accessible chromatin-photoactivated localization microscopy (3D ATAC-PALM) that integrates an assay for transposase-accessible chromatin with visualization, PALM super-resolution imaging and lattice light-sheet microscopy. Multiplexed with oligopaint DNA–fluorescence in situ hybridization (FISH), RNA–FISH and protein fluorescence, 3D ATAC-PALM connected microscopy and genomic data, revealing spatially segregated accessible chromatin domains (ACDs) that enclose active chromatin and transcribed genes. Using these methods to analyze genetically perturbed cells, we demonstrated that genome architectural protein CTCF prevents excessive clustering of accessible chromatin and decompacts ACDs. These results highlight 3D ATAC-PALM as a useful tool to probe the structure and organizing mechanism of the genome.
Single-molecule localisation microscopy (SMLM) allows the localisation of fluorophores with a precision of 10-30 nm, revealing the cell's nanoscale architecture at the molecular level. Recently, SMLM has been extended to 3D, providing a unique insight into cellular machinery. Although cluster analysis techniques have been developed for 2D SMLM data sets, few have been applied to 3D. This lack of quantification tools can be explained by the relative novelty of imaging techniques such as interferometric photo-activated localisation microscopy (iPALM). Also, existing methods that could be extended to 3D SMLM are usually subject to user defined analysis parameters, which remains a major drawback. Here, we present a new open source cluster analysis method for 3D SMLM data, free of user definable parameters, relying on a model-based Bayesian approach which takes full account of the individual localisation precisions in all three dimensions. The accuracy and reliability of the method is validated using simulated data sets. This tool is then deployed on novel experimental data as a proof of concept, illustrating the recruitment of LAT to the T-cell immunological synapse in data acquired by iPALM providing ~10 nm isotropic resolution.
Microtubules play a major role in intracellular trafficking of vesicles in endocrine cells. Detailed knowledge of microtubule organization and their relation to other cell constituents is crucial for understanding cell function. However, their role in insulin transport and secretion is under debate. Here, we use FIB-SEM to image islet β cells in their entirety with unprecedented resolution. We reconstruct mitochondria, Golgi apparati, centrioles, insulin secretory granules, and microtubules of seven β cells, and generate a comprehensive spatial map of microtubule-organelle interactions. We find that microtubules form nonradial networks that are predominantly not connected to either centrioles or endomembranes. Microtubule number and length, but not microtubule polymer density, vary with glucose stimulation. Furthermore, insulin secretory granules are enriched near the plasma membrane, where they associate with microtubules. In summary, we provide the first 3D reconstructions of complete microtubule networks in primary mammalian cells together with evidence regarding their importance for insulin secretory granule positioning and thus their supportive role in insulin secretion.

Object detection and classification are key tasks in computer vision that can facilitate high-throughput image analysis of microscopy data. We present a set of local image descriptors for three-dimensional (3D) microscopy datasets inspired by the well-known Haar wavelet framework. We add orientation, illumination and scale information by assuming that the neighborhood surrounding points of interests in the image can be described with ellipsoids, and we increase discriminative power by incorporating edge and shape information into the features. The calculation of the local image descriptors is implemented in a Graphics Processing Unit (GPU) in order to reduce computation time to 1 millisecond per object of interest. We present results for cell division detection in 3D time-lapse fluorescence microscopy with 97.6% accuracy.
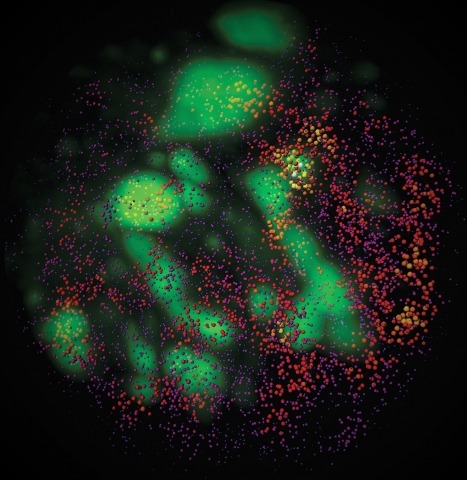
Combinatorial cis-regulatory networks encoded in animal genomes represent the foundational gene expression mechanism for directing cell-fate commitment and maintenance of cell identity by transcription factors (TFs). However, the 3D spatial organization of cis-elements and how such sub-nuclear structures influence TF activity remain poorly understood. Here, we combine lattice light-sheet imaging, single-molecule tracking, numerical simulations, and ChIP-exo mapping to localize and functionally probe Sox2 enhancer-organization in living embryonic stem cells. Sox2 enhancers form 3D-clusters that are segregated from heterochromatin but overlap with a subset of Pol II enriched regions. Sox2 searches for specific binding targets via a 3D-diffusion dominant mode when shuttling long-distances between clusters while chromatin-bound states predominate within individual clusters. Thus, enhancer clustering may reduce global search efficiency but enables rapid local fine-tuning of TF search parameters. Our results suggest an integrated model linking cis-element 3D spatial distribution to local-versus-global target search modalities essential for regulating eukaryotic gene transcription.
3D live imaging is important for a better understanding of biological processes, but it is challenging with current techniques such as spinning-disk confocal microscopy. Bessel beam plane illumination microscopy allows high-speed 3D live fluorescence imaging of living cellular and multicellular specimens with nearly isotropic spatial resolution, low photobleaching and low photodamage. Unlike conventional fluorescence imaging techniques that usually have a unique operation mode, Bessel plane illumination has several modes that offer different performance with different imaging metrics. To achieve optimal results from this technique, the appropriate operation mode needs to be selected and the experimental setting must be optimized for the specific application and associated sample properties. Here we explain the fundamental working principles of this technique, discuss the pros and cons of each operational mode and show through examples how to optimize experimental parameters. We also describe the procedures needed to construct, align and operate a Bessel beam plane illumination microscope by using our previously reported system as an example, and we list the necessary equipment to build such a microscope. Assuming all components are readily available, it would take a person skilled in optical instrumentation \~{}1 month to assemble and operate a microscope according to this protocol.