Filter
Result Type
Associated Lab
- Aso Lab (1) Apply Aso Lab filter
- Baker Lab (3) Apply Baker Lab filter
- Betzig Lab (7) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Cui Lab (3) Apply Cui Lab filter
- Dickson Lab (1) Apply Dickson Lab filter
- Dudman Lab (1) Apply Dudman Lab filter
- Eddy/Rivas Lab (5) Apply Eddy/Rivas Lab filter
- Fetter Lab (2) Apply Fetter Lab filter
- Gonen Lab (1) Apply Gonen Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Keller Lab (2) Apply Keller Lab filter
- Lavis Lab (5) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Rubin Lab (3) Apply Rubin Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (2) Apply Schreiter Lab filter
- Simpson Lab (3) Apply Simpson Lab filter
- Singer Lab (1) Apply Singer Lab filter
- Sternson Lab (6) Apply Sternson Lab filter
- Svoboda Lab (7) Apply Svoboda Lab filter
- Tjian Lab (2) Apply Tjian Lab filter
- Truman Lab (1) Apply Truman Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
- Zuker Lab (2) Apply Zuker Lab filter
Associated Project Team
Publication Date
- December 2011 (10) Apply December 2011 filter
- November 2011 (8) Apply November 2011 filter
- October 2011 (8) Apply October 2011 filter
- September 2011 (8) Apply September 2011 filter
- August 2011 (9) Apply August 2011 filter
- July 2011 (5) Apply July 2011 filter
- June 2011 (10) Apply June 2011 filter
- May 2011 (6) Apply May 2011 filter
- April 2011 (5) Apply April 2011 filter
- March 2011 (6) Apply March 2011 filter
- February 2011 (8) Apply February 2011 filter
- January 2011 (15) Apply January 2011 filter
- Remove 2011 filter 2011
98 Results
Showing 1-10 of 98 resultsLongitudinal axon fascicles within the Drosophila embryonic CNS provide connections between body segments and are required for coordinated neural signaling along the anterior-posterior axis. We show here that establishment of select CNS longitudinal tracts and formation of precise mechanosensory afferent innervation to the same CNS region are coordinately regulated by the secreted semaphorins Sema-2a and Sema-2b. Both Sema-2a and Sema-2b utilize the same neuronal receptor, plexin B (PlexB), but serve distinct guidance functions. Localized Sema-2b attraction promotes the initial assembly of a subset of CNS longitudinal projections and subsequent targeting of chordotonal sensory afferent axons to these same longitudinal connectives, whereas broader Sema-2a repulsion serves to prevent aberrant innervation. In the absence of Sema-2b or PlexB, chordotonal afferent connectivity within the CNS is severely disrupted, resulting in specific larval behavioral deficits. These results reveal that distinct semaphorin-mediated guidance functions converge at PlexB and are critical for functional neural circuit assembly.

A crucial issue in studies of morphogen gradients relates to their range: the distance over which they can act as direct regulators of cell signaling, gene expression and cell differentiation. To address this, we present a straightforward statistical framework that can be used in multiple developmental systems. We illustrate the developed approach by providing a point estimate and confidence interval for the spatial range of the graded distribution of nuclear Dorsal, a transcription factor that controls the dorsoventral pattern of the Drosophila embryo.
Mitochondria are highly dynamic organelles that play multiple roles in cells. How mitochondria cooperatively modulate embryonic stem (ES) cell function during development is not fully understood. Global disruption of Ptpmt1, a mitochondrial Pten-like phosphatidylinositol phosphate (PIP) phosphatase, resulted in developmental arrest and postimplantation lethality. Ptpmt1(-/-) blastocysts failed to outgrow, and inner-cell-mass cells failed to thrive. Depletion of Ptpmt1 in conditional knockout ES cells decreased proliferation without affecting energy homeostasis or cell survival. Differentiation of Ptpmt1-depleted ES cells was essentially blocked. This was accompanied by upregulation of cyclin-dependent kinase inhibitors and a significant cell cycle delay. Reintroduction of wild-type but not of catalytically deficient Ptpmt1 C132S or truncated Ptpmt1 lacking the mitochondrial localization signal restored the differentiation capabilities of Ptpmt1 knockout ES cells. Intriguingly, Ptpmt1 is specifically important for stem cells, as ablation of Ptpmt1 in differentiated embryonic fibroblasts did not disturb cellular function. Further analyses demonstrated that oxygen consumption of Ptpmt1-depleted cells was decreased, while glycolysis was concomitantly enhanced. In addition, mitochondrial fusion/dynamics were compromised in Ptpmt1 knockout cells due to accumulation of PIPs. These studies, while establishing a crucial role for Ptpmt1 phosphatase in embryogenesis, reveal a mitochondrial metabolic stress-activated checkpoint in the control of ES cell differentiation.
We describe the generation of a family of high-signal-to-noise single-wavelength genetically encoded indicators for maltose. This was achieved by insertion of circularly permuted fluorescent proteins into a bacterial periplasmic binding protein (PBP), Escherichia coli maltodextrin-binding protein, resulting in a four-color family of maltose indicators. The sensors were iteratively optimized to have sufficient brightness and maltose-dependent fluorescence increases for imaging, under both one- and two-photon illumination. We demonstrate that maltose affinity of the sensors can be tuned in a fashion largely independent of the fluorescent readout mechanism. Using literature mutations, the binding specificity could be altered to moderate sucrose preference, but with a significant loss of affinity. We use the soluble sensors in individual E. coli bacteria to observe rapid maltose transport across the plasma membrane, and membrane fusion versions of the sensors on mammalian cells to visualize the addition of maltose to extracellular media. The PBP superfamily includes scaffolds specific for a number of analytes whose visualization would be critical to the reverse engineering of complex systems such as neural networks, biosynthetic pathways, and signal transduction cascades. We expect the methodology outlined here to be useful in the development of indicators for many such analytes.
A large number of degrees of freedom are required to produce a high quality focus through random scattering media. Previous demonstrations based on spatial phase modulations suffer from either a slow speed or a small number of degrees of freedom. In this work, a high speed wavefront determination technique based on spatial frequency domain wavefront modulations is proposed and experimentally demonstrated, which is capable of providing both a high operation speed and a large number of degrees of freedom. The technique was employed to focus light through a strongly scattering medium and the entire wavefront was determined in 400 milliseconds, three orders of magnitude faster than the previous report.
Profile hidden Markov models (profile HMMs) and probabilistic inference methods have made important contributions to the theory of sequence database homology search. However, practical use of profile HMM methods has been hindered by the computational expense of existing software implementations. Here I describe an acceleration heuristic for profile HMMs, the "multiple segment Viterbi" (MSV) algorithm. The MSV algorithm computes an optimal sum of multiple ungapped local alignment segments using a striped vector-parallel approach previously described for fast Smith/Waterman alignment. MSV scores follow the same statistical distribution as gapped optimal local alignment scores, allowing rapid evaluation of significance of an MSV score and thus facilitating its use as a heuristic filter. I also describe a 20-fold acceleration of the standard profile HMM Forward/Backward algorithms using a method I call "sparse rescaling". These methods are assembled in a pipeline in which high-scoring MSV hits are passed on for reanalysis with the full HMM Forward/Backward algorithm. This accelerated pipeline is implemented in the freely available HMMER3 software package. Performance benchmarks show that the use of the heuristic MSV filter sacrifices negligible sensitivity compared to unaccelerated profile HMM searches. HMMER3 is substantially more sensitive and 100- to 1000-fold faster than HMMER2. HMMER3 is now about as fast as BLAST for protein searches.
Understanding the structure and function of neural circuits are central questions in neuroscience research. To address these questions, new genetically encoded tools have been developed for mapping, monitoring, and manipulating neurons. Essential to implementation of these tools is their selective delivery to defined neuronal populations in the brain. This has been facilitated by recent improvements in cell type-specific transgene expression using recombinant adeno-associated viral vectors. Here, we highlight these developments and discuss areas for improvement that could further expand capabilities for neural circuit analysis.
Electron crystallography is a powerful technique for the study of membrane protein structure and function in the lipid environment. When well-ordered two-dimensional crystals are obtained the structure of both protein and lipid can be determined and lipid-protein interactions analyzed. Protons and ionic charges can be visualized by electron crystallography and the protein of interest can be captured for structural analysis in a variety of physiologically distinct states. This review highlights the strengths of electron crystallography and the momentum that is building up in automation and the development of high throughput tools and methods for structural and functional analysis of membrane proteins by electron crystallography.
Small molecule fluorophores are essential tools for chemical biology. A benefit of synthetic dyes is the ability to employ chemical approaches to control the properties and direct the position of the fluorophore. Applying modern synthetic organic chemistry strategies enables efficient tailoring of the chemical structure to obtain probes for specific biological experiments. Chemistry can also be used to activate fluorophores; new fluorogenic enzyme substrates and photoactivatable compounds with improved properties have been prepared that facilitate advanced imaging experiments with low background fluorescence. Finally, chemical reactions in live cells can be used to direct the spatial distribution of the fluorophore, allowing labeling of defined cellular regions with synthetic dyes.
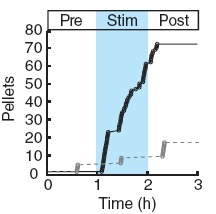
Two intermingled hypothalamic neuron populations specified by expression of agouti-related peptide (AGRP) or pro-opiomelanocortin (POMC) positively and negatively influence feeding behavior, respectively, possibly by reciprocally regulating downstream melanocortin receptors. However, the sufficiency of these neurons to control behavior and the relationship of their activity to the magnitude and dynamics of feeding are unknown. To measure this, we used channelrhodopsin-2 for cell type-specific photostimulation. Activation of only 800 AGRP neurons in mice evoked voracious feeding within minutes. The behavioral response increased with photoexcitable neuron number, photostimulation frequency and stimulus duration. Conversely, POMC neuron stimulation reduced food intake and body weight, which required melanocortin receptor signaling. However, AGRP neuron-mediated feeding was not dependent on suppressing this melanocortin pathway, indicating that AGRP neurons directly engage feeding circuits. Furthermore, feeding was evoked selectively over drinking without training or prior photostimulus exposure, which suggests that AGRP neurons serve a dedicated role coordinating this complex behavior.