Filter
Associated Lab
- Aguilera Castrejon Lab (16) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (64) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (112) Apply Betzig Lab filter
- Beyene Lab (13) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (52) Apply Branson Lab filter
- Card Lab (40) Apply Card Lab filter
- Cardona Lab (63) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (50) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (19) Apply Espinosa Medina Lab filter
- Feliciano Lab (7) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- Fitzgerald Lab (29) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (38) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (60) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (26) Apply Hermundstad Lab filter
- Hess Lab (76) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (46) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (6) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (76) Apply Keller Lab filter
- Koay Lab (18) Apply Koay Lab filter
- Lavis Lab (148) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (27) Apply Li Lab filter
- Lippincott-Schwartz Lab (167) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (6) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (61) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (46) Apply Pachitariu Lab filter
- Pastalkova Lab (18) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (51) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (43) Apply Romani Lab filter
- Rubin Lab (143) Apply Rubin Lab filter
- Saalfeld Lab (62) Apply Saalfeld Lab filter
- Satou Lab (16) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (67) Apply Schreiter Lab filter
- Sgro Lab (20) Apply Sgro Lab filter
- Shroff Lab (29) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (93) Apply Spruston Lab filter
- Stern Lab (156) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (33) Apply Stringer Lab filter
- Svoboda Lab (135) Apply Svoboda Lab filter
- Tebo Lab (33) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (21) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (49) Apply Turaga Lab filter
- Turner Lab (37) Apply Turner Lab filter
- Vale Lab (7) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (17) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (2) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (10) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (53) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (45) Apply GENIE filter
- Integrative Imaging (2) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (26) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2025 (72) Apply 2025 filter
- 2024 (223) Apply 2024 filter
- 2023 (163) Apply 2023 filter
- 2022 (193) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4064 Publications
Showing 4051-4060 of 4064 resultsCell fate choice is a key event happening during preimplantation mouse development. From embryonic day 3.5 (E3.5) to E4.5, the inner cell mass (ICM) differentiates into epiblast (Epi, NANOG expressing cells) and primitive endoderm (PrE, GATA6, SOX17 and/or GATA4 expressing cells). The mechanism by which ICM cells differentiate into Epi cells and PrE cells remains partially unknown. FGF/ERK has been proposed as the main signalling pathway for this event, but it does not explain co-expression of NANOG and GAT6 or how the cell fate choice is initiated. In this study, we investigate whether Wnt/β-catenin signalling also plays a role. To this end, we use two in vitro models based on inducible GATA6 expression: one in 2D, and another in 3D, namely ICM organoids. By combining these in vitro models with in vivo mouse embryos, chemical and classical genetics, and quantitative 3D immunofluorescence analyses, we propose a dual role for Wnt/β-catenin signalling. We find that β-catenin, acting alongside FGF/ERK signalling, helps to guide the cell fate choice towards PrE. Additionally, by regulating GATA6 and GATA4 stability, β-catenin further facilitates this choice. To summarise, we observe that pathway activation promotes PrE differentiation, while its inhibition stalls it. SUMMARY STATEMENT Wnt/β-catenin signalling promotes PrE fate in mouse preimplantation embryos.
In serial recall experiments, human subjects are requested to retrieve a list of words in the same order as they were presented. In a classical study, participants were reported to recall more words from study lists composed of short words compared to lists of long words, the word length effect. The world length effect was also observed in free recall experiments, where subjects can retrieve the words in any order. Here we analyzed a large dataset from free recall experiments of unrelated words, where short and long words were randomly mixed, and found a seemingly opposite effect: long words are recalled better than the short ones. We show that our recently proposed mechanism of associative retrieval can explain both these observations. Moreover, the direction of the effect depends solely on the way study lists are composed.
It has been proposed that dosage compensation in Drosophila males occurs by binding of two core proteins, MSL-1 and MSL-2, to a set of 35-40 X chromosome "entry sites" that serve to nucleate mature complexes, termed compensasomes, which then spread to neighboring sequences to double expression of most X-linked genes. Here we show that any piece of the X chromosome with which compensasomes are associated in wild-type displays a normal pattern of compensasome binding when inserted into an autosome, independently of the presence of an entry site. Furthermore, in chromosomal rearrangements in which a piece of X chromosome is inserted into an autosome, or a piece of autosome is translocated to the X chromosome, we do not observe spreading of compensasomes to regions of autosomes that have been juxtaposed to X chromosomal material. Taken together these results suggest that spreading is not involved in dosage compensation and that nothing distinguishes an entry site from the other X chromosome sites occupied by compensasomes beyond their relative affinities for compensasomes. We propose a new model in which the distribution of compensasomes along the X chromosome is achieved according to the hierarchical affinities of individual binding sites.
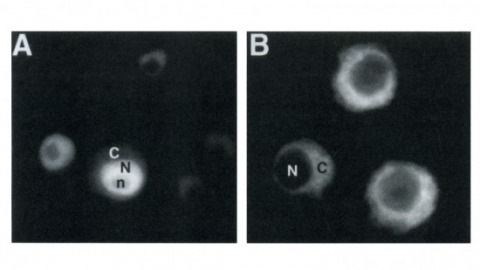
Drosophila yan has been postulated to act as an antagonist of the proneural signal mediated by the sevenless/Ras1/MAPK pathway. We have mutagenized the eight MAPK phosphorylation consensus sites of yan and examined the effects of overexpressing the mutant protein in transgenic flies and transfected S2 cultured cells. Our results suggest that phosphorylation by MAPK affects the stability and subcellular localization of yan, resulting in rapid down-regulation of yan activity. Furthermore, MAPK-mediated down-regulation of yan function appears to be critical for the proper differentiation of both neuronal and nonneuronal tissues throughout development, suggesting that yan is an essential component of a general timing mechanism controlling the competence of a cell to respond to inductive signals.
YAP/TEAD signaling is essential for organismal development, cell proliferation, and cancer progression. As a transcriptional coactivator, how YAP activates its downstream target genes is incompletely understood. YAP forms biomolecular condensates in response to hyperosmotic stress, concentrating transcription-related factors to activate downstream target genes. However, whether YAP forms condensates under other signals, how YAP condensates organize and function, and how YAP condensates activate transcription in general are unknown. Here, we report that endogenous YAP forms sub-micron scale condensates in response to Hippo pathway regulation and actin cytoskeletal tension. YAP condensates are stabilized by the transcription factor TEAD1, and recruit BRD4, a coactivator that is enriched at active enhancers. Using single-particle tracking, we found that YAP condensates slowed YAP diffusion within condensate boundaries, a possible mechanism for promoting YAP target search. These results reveal that YAP condensate formation is a highly regulated process that is critical for YAP/TEAD target gene expression.
Cells in many tissues, such as bone, muscle, and placenta, fuse into syncytia to acquire new functions and transcriptional programs. While it is known that fused cells are specialized, it is unclear whether cell-fusion itself contributes to programmatic-changes that generate the new cellular state. Here, we address this by employing a fusogen-mediated, cell-fusion system to create syncytia from undifferentiated cells. RNA-Seq analysis reveals VSV-G-induced cell fusion precedes transcriptional changes. To gain mechanistic insights, we measure the plasma membrane surface area after cell-fusion and observe it diminishes through increases in endocytosis. Consequently, glucose transporters internalize, and cytoplasmic glucose and ATP transiently decrease. This reduced energetic state activates AMPK, which inhibits YAP1, causing transcriptional-reprogramming and cell-cycle arrest. Impairing either endocytosis or AMPK activity prevents YAP1 inhibition and cell-cycle arrest after fusion. Together, these data demonstrate plasma membrane diminishment upon cell-fusion causes transient nutrient stress that may promote transcriptional-reprogramming independent from extrinsic cues.
Analysis of the nucleotide sequence of the genetic locus for yeast mitochondrial RNA polymerase (RPO41) reveals a continuous open reading frame with the coding potential for a polypeptide of 1351 amino acids, a size consistent with the electrophoretic mobility of this enzymatic activity. The transcription product from this gene spans the singular reading frame. In vivo transcript abundance reflects codon usage and growth under stringent conditions for mitochondrial biogenesis and function results in a several fold higher level of gene expression than growth under glucose repression. A comparison of the yeast mitochondrial RNA polymerase amino acid sequence to those of E. coli RNA polymerase subunits failed to demonstrate any regions of homology. Interestingly, the mitochondrial enzyme is highly homologous to the DNA-directed RNA polymerases of bacteriophages T3 and T7, especially in regions most highly conserved between the T3 and T7 enzymes themselves.
The vast majority of mammalian genomes are transcribed as non-coding RNA in what is referred to as “pervasive transcription.” Recent studies have uncovered various families of non-coding RNA transcribed upstream of transcription start sites. In particular, highly unstable promoter upstream transcripts known as PROMPTs have been shown to be targeted for exosomal degradation by the nuclear exosome targeting complex (NEXT) consisting of the RNA helicase MTR4, the zinc-knuckle scaffold ZCCHC8, and the RNA binding protein RBM7. Here, we report that in addition to its known RNA substrates, ZCCHC8 is required for the targeted degradation of pervasive transcripts produced at CTCF binding sites, open chromatin regions, promoters, promoter flanking regions, and transcription factor binding sites. Additionally, we report that a significant number of RIKEN cDNAs and predicted genes display the hallmarks of PROMPTs and are also substrates for ZCCHC8 and/or NEXT complex regulation suggesting these are unlikely to be functional genes. Our results suggest that ZCCHC8 and/or the NEXT complex may play a larger role in the global regulation of pervasive transcription than previously reported.Competing Interest StatementThe authors have declared no competing interest.
Brains are notoriously hard to understand, and neuroscientists need all the tools they can get their hands on to have a realistic shot at it. Advances in machine learning are proving instrumental, illustrated by their recent use to shed light on navigational strategies implemented by zebrafish brains.