Filter
Associated Lab
- Aguilera Castrejon Lab (19) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (73) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Beyene Lab (15) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (55) Apply Branson Lab filter
- Card Lab (43) Apply Card Lab filter
- Cardona Lab (64) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (16) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (3) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (56) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (23) Apply Espinosa Medina Lab filter
- Feliciano Lab (12) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (30) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (65) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (30) Apply Hermundstad Lab filter
- Hess Lab (80) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (48) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (6) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (76) Apply Keller Lab filter
- Koay Lab (19) Apply Koay Lab filter
- Lavis Lab (158) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (32) Apply Li Lab filter
- Lippincott-Schwartz Lab (180) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (8) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (65) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (8) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (52) Apply Pachitariu Lab filter
- Pastalkova Lab (19) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (55) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (51) Apply Romani Lab filter
- Rubin Lab (149) Apply Rubin Lab filter
- Saalfeld Lab (64) Apply Saalfeld Lab filter
- Satou Lab (18) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (70) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (23) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (98) Apply Spruston Lab filter
- Stern Lab (160) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (41) Apply Stringer Lab filter
- Svoboda Lab (136) Apply Svoboda Lab filter
- Tebo Lab (35) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (53) Apply Turaga Lab filter
- Turner Lab (38) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (4) Apply Voigts Lab filter
- Wang (Meng) Lab (29) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (9) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2026 (70) Apply 2026 filter
- 2025 (222) Apply 2025 filter
- 2024 (210) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (192) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4265 Publications
Showing 1701-1710 of 4265 resultsDrosophila sensory neurons form distinctive terminal branch patterns in the developing neuropile of the embryonic central nervous system. In this paper we make a genetic analysis of factors regulating arbor position. We show that mediolateral position is determined in a binary fashion by expression (chordotonal neurons) or nonexpression (multidendritic neurons) of the Robo3 receptor for the midline repellent Slit. Robo3 expression is one of a suite of chordotonal neuron properties that depend on expression of the proneural gene atonal. Different features of terminal branches are separately regulated: an arbor can be shifted mediolaterally without affecting its dorsoventral location, and the distinctive remodeling of one arbor continues as normal despite this arbor shifting to an abnormal position in the neuropile.
The insect juvenile hormone receptor is a basic helix-loop-helix (bHLH), Per-Arnt-Sim (PAS) domain protein, a novel type of hormone receptor. In higher flies like Drosophila, the ancestral receptor germ cell-expressed (gce) gene has duplicated to yield the paralog Methoprene-tolerant (Met). These paralogous receptors share redundant function during development but play unique roles in adults. Some aspects of JH function apparently require one receptor or the other. To provide a foundation for studying JH receptor function, we have recapitulated endogenous JH receptor expression with single cell resolution. Using Bacteria Artificial Chromosome (BAC) recombineering and a transgenic knock-in, we have generated a spatiotemporal expressional atlas of Metand gce throughout development. We demonstrate JH receptor expression in known JH target tissues, in which temporal expression corresponds with periods of hormone sensitivity. Larval expression largely supports the notion of functional redundancy. Furthermore, we provide the neuroanatomical distribution of JH receptors in both the larval and adult central nervous system, which will serve as a platform for future studies regarding JH action on insect behavior.
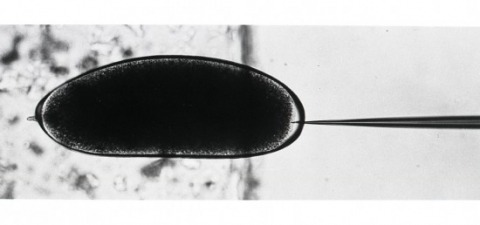
Exogenous DNA sequences were introduced into the Drosophila germ line. A rosy transposon (ry1), constructed by inserting a chromosomal DNA fragment containing the wild-type rosy gene into a P transposable element, transformed germ line cells in 20 to 50 percent of the injected rosy mutant embryos. Transformants contained one or two copies of chromosomally integrated, intact ry1 that were stably inherited in subsequent generations. These transformed flies had wild-type eye color indicating that the visible genetic defect in the host strain could be fully and permanently corrected by the transferred gene. To demonstrate the generality of this approach, a DNA segment that does not confer a recognizable phenotype on recipients was also transferred into germ line chromosomes.
Many polyphenisms are examples of adaptive phenotypic plasticity where a single genotype produces distinct phenotypes in response to environmental cues. Such alternative phenotypes occur as winged and wingless parthenogenetic females in the pea aphid (Acyrthosiphon pisum). However, the proportion of winged females produced in response to a given environmental cue varies between clonal genotypes. Winged and wingless phenotypes also occur in males of the sexual generation. In contrast to parthenogenetic females, wing production in males is environmentally insensitive and controlled by the sex-linked, biallelic locus, aphicarus (api). Hence, environmental or genetic cues induce development of winged and wingless phenotypes at different stages of the pea aphid life cycle. We have tested whether allelic variation at the api locus explains genetic variation in the propensity to produce winged females. We assayed clones from an F2 cross that were heterozygous or homozygous for alternative api alleles for their propensity to produce winged offspring. We found that clones with different api genotypes differed in their propensity to produce winged offspring. The results indicate genetic linkage of factors controlling the female wing polyphenism and male wing polymorphism. This finding is consistent with the hypothesis that genotype by environment interaction at the api locus explains genetic variation in the environmentally cued wing polyphenism.
The present disclosure provides, inter alia, genetically encoded recombinant peptide biosensors comprising analyte-binding framework portions and signaling portions, wherein the signaling portions are present within the framework portions at sites or amino acid positions that undergo a conformational change upon interaction of the framework portion with an analyte.
The discovery of intracellular Ca(2+) signals within astrocytes has changed our view of how these ubiquitous cells contribute to brain function. Classically thought merely to serve supportive functions, astrocytes are increasingly thought to respond to, and regulate, neurons. The use of organic Ca(2+) indicator dyes such as Fluo-4 and Fura-2 has proved instrumental in the study of astrocyte physiology. However, progress has recently been accelerated by the use of cytosolic and membrane targeted genetically encoded calcium indicators (GECIs). Herein, we review these recent findings, discuss why studying astrocyte Ca(2+) signals is important to understand brain function, and summarize work that led to the discovery of TRPA1 channel-mediated near-membrane Ca(2+) signals in astrocytes and their indirect neuromodulatory roles at inhibitory synapses in the CA1 stratum radiatum region of the hippocampus. We suggest that the use of membrane-targeted and cytosolic GECIs holds great promise to explore the diversity of Ca(2+) signals within single astrocytes and also to study diversity of function for astrocytes in different parts of the brain.
Genetically encoded calcium indicators (GECIs) are powerful tools for systems neuroscience. Here we describe red, single-wavelength GECIs, "RCaMPs," engineered from circular permutation of the thermostable red fluorescent protein mRuby. High-resolution crystal structures of mRuby, the red sensor RCaMP, and the recently published red GECI R-GECO1 give insight into the chromophore environments of the Ca(2+)-bound state of the sensors and the engineered protein domain interfaces of the different indicators. We characterized the biophysical properties and performance of RCaMP sensors in vitro and in vivo in Caenorhabditis elegans, Drosophila larvae, and larval zebrafish. Further, we demonstrate 2-color calcium imaging both within the same cell (registering mitochondrial and somatic [Ca(2+)]) and between two populations of cells: neurons and astrocytes. Finally, we perform integrated optogenetics experiments, wherein neural activation via channelrhodopsin-2 (ChR2) or a red-shifted variant, and activity imaging via RCaMP or GCaMP, are conducted simultaneously, with the ChR2/RCaMP pair providing independently addressable spectral channels. Using this paradigm, we measure calcium responses of naturalistic and ChR2-evoked muscle contractions in vivo in crawling C. elegans. We systematically compare the RCaMP sensors to R-GECO1, in terms of action potential-evoked fluorescence increases in neurons, photobleaching, and photoswitching. R-GECO1 displays higher Ca(2+) affinity and larger dynamic range than RCaMP, but exhibits significant photoactivation with blue and green light, suggesting that integrated channelrhodopsin-based optogenetics using R-GECO1 may be subject to artifact. Finally, we create and test blue, cyan, and yellow variants engineered from GCaMP by rational design. This engineered set of chromatic variants facilitates new experiments in functional imaging and optogenetics.
Neurons and glia are functionally organized into circuits and higher-order structures via synaptic connectivity, well-orchestrated molecular signaling, and activity-dependent refinement. Such organization allows the precise information processing required for complex behaviors. Disruption of nervous systems by genetic deficiency or events such as trauma or environmental exposure may produce a diseased state in which certain aspects of inter-neuron signaling are impaired. Optical imaging techniques allow the direct visualization of individual neurons in a circuit environment. Imaging probes specific for given biomolecules may help elucidate their contribution to proper circuit function. Genetically encoded sensors can visualize trafficking of particular molecules in defined neuronal populations, non-invasively in intact brain or reduced preparations. Sensor analysis in healthy and diseased brains may reveal important differences and shed light on the development and progression of nervous system disorders. We review the field of genetically encoded sensors for molecules and cellular events, and their potential applicability to the study of nervous system disease.
Recording activity from identified populations of neurons is a central goal of neuroscience. Changes in membrane depolarization, particularly action potentials, are the most important features of neural physiology to extract, although ions, neurotransmitters, neuromodulators, second messengers, and the activation state of specific proteins are also crucial. Modern fluorescence microscopy provides the basis for such activity mapping, through multi-photon imaging and other optical schemes. Probes remain the rate-limiting step for progress in this field: they should be bright and photostable, and ideally come in multiple colors. Only protein-based reagents permit chronic imaging from genetically specified cells. Here we review recent progress in the design, optimization and deployment of genetically encoded indicators for calcium ions (a proxy for action potentials), membrane potential, and neurotransmitters. We highlight seminal experiments, and present an outlook for future progress.
The basolateral amygdala (BLA) plays essential roles in behaviors motivated by stimuli with either positive or negative valence, but how it processes motivationally opposing information and participates in establishing valence-specific behaviors remains unclear. Here, by targeting Fezf2-expressing neurons in the BLA, we identify and characterize two functionally distinct classes in behaving mice, the negative-valence neurons and positive-valence neurons, which innately represent aversive and rewarding stimuli, respectively, and through learning acquire predictive responses that are essential for punishment avoidance or reward seeking. Notably, these two classes of neurons receive inputs from separate sets of sensory and limbic areas, and convey punishment and reward information through projections to the nucleus accumbens and olfactory tubercle, respectively, to drive negative and positive reinforcement. Thus, valence-specific BLA neurons are wired with distinctive input-output structures, forming a circuit framework that supports the roles of the BLA in encoding, learning and executing valence-specific motivated behaviors.