Filter
Associated Lab
- Aguilera Castrejon Lab (19) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (75) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Beyene Lab (15) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (56) Apply Branson Lab filter
- Card Lab (43) Apply Card Lab filter
- Cardona Lab (64) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (16) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (3) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (58) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (25) Apply Espinosa Medina Lab filter
- Feliciano Lab (16) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (30) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (65) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (32) Apply Hermundstad Lab filter
- Hess Lab (80) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (49) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (7) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (15) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (77) Apply Keller Lab filter
- Koay Lab (20) Apply Koay Lab filter
- Lavis Lab (162) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (32) Apply Li Lab filter
- Lippincott-Schwartz Lab (182) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (9) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (65) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (8) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (56) Apply Pachitariu Lab filter
- Pastalkova Lab (19) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (55) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (52) Apply Romani Lab filter
- Rubin Lab (149) Apply Rubin Lab filter
- Saalfeld Lab (66) Apply Saalfeld Lab filter
- Satou Lab (18) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (72) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (23) Apply Sgro Lab filter
- Shroff Lab (35) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (98) Apply Spruston Lab filter
- Stern Lab (160) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (44) Apply Stringer Lab filter
- Svoboda Lab (136) Apply Svoboda Lab filter
- Tebo Lab (36) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (53) Apply Turaga Lab filter
- Turner Lab (38) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (5) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (11) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2026 (118) Apply 2026 filter
- 2025 (223) Apply 2025 filter
- 2024 (209) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (192) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4313 Publications
Showing 2131-2140 of 4313 resultsTransfer of autoantibodies targeting ionotropic N-methyl-D-aspartate receptors in autoimmune encephalitis patients into mice leads to typical disease signs. Long-term effects of the pathogenic antibodies consist of immunoglobulin G-induced crosslinking and receptor internalization. We focused on the direct and immediate impact of a specific pathogenic patient-derived monoclonal autoantibody (immunoglobulin G #003-102) on receptor function.We performed cell-attached recordings in cells transfected with the GluN1 and GluN2A subunit of the N-methyl-D-aspartate receptor. Immunoglobulin G #003-102 binds to the amino-terminal domain of the glycine-binding GluN1 subunit. It reduced simultaneous receptor openings significantly compared to controls at both low and high glutamate and glycine concentrations. Closer examination of our data in 50-second to 2-second intervals revealed, that Immunoglobulin G #003-102 rapidly decreases the number of open receptors. However, antigen-binding fragments of immunoglobulin G #003-102 did not reduce the receptor openings.In conclusion, patient-derived immunoglobulin G #003-102 inhibits N-methyl-D-aspartate receptors rapidly and directly before receptor internalization occurs and the entire immunoglobulin G is necessary for this acute inhibitory effect. This suggests an application of the antigen-binding fragment-like constructs of #003-102 as a potential new treatment strategy for shielding the pathogenic epitopes on the N-methyl-D-aspartate receptors.
An automated ultra-microtome capable of sectioning thousands of ultrathin sections onto standard TEM slot grids was developed and used to section: a complete Drosophila melanogaster first-instar larva, three sections per grid, into 4,866 34-nm-thick sections with a cutting and pickup success rate of 99.74%; 30 microns of mouse cortex measuring roughly 400 um x 2000 um at 40 nm per slice; and a full adult Drosophila brain and ventral nerve column into 9,300 sections with a pickup success rate of 99.95%. The apparatus uses optical interferometers to monitor a reference distance between the cutting knife and multiple sample blocks. Cut sections are picked up from the knife-boat water surface while they are still anchored to the cutting knife. Blocks without embedded tissue are used to displace tissue-containing sections away from the knife edge so that the tissue regions end up in the grid slot instead of on the grid rim.
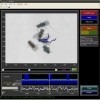
We present a machine learning–based system for automatically computing interpretable, quantitative measures of animal behavior. Through our interactive system, users encode their intuition about behavior by annotating a small set of video frames. These manual labels are converted into classifiers that can automatically annotate behaviors in screen-scale data sets. Our general-purpose system can create a variety of accurate individual and social behavior classifiers for different organisms, including mice and adult and larval Drosophila.
Janelia Farm, the new research campus of the Howard Hughes Medical Institute, is an ongoing experiment in the social engineering of research communities.
Neuronal signal transduction by the JNK MAP kinase pathway is altered by a broad array of stimuli including exposure to the widely abused drug ethanol, but the behavioral relevance and the regulation of JNK signaling is unclear. Here we demonstrate that JNK signaling functions downstream of the Sterile20 kinase family gene tao/Taok3 to regulate the behavioral effects of acute ethanol exposure in both the fruit fly Drosophila and mice. In flies tao is required in neurons to promote sensitivity to the locomotor stimulant effects of acute ethanol exposure and to establish specific brain structures. Reduced expression of key JNK pathway genes substantially rescued the structural and behavioral phenotypes of tao mutants. Decreasing and increasing JNK pathway activity resulted in increased and decreased sensitivity to the locomotor stimulant properties of acute ethanol exposure, respectively. Further, JNK expression in a limited pattern of neurons that included brain regions implicated in ethanol responses was sufficient to restore normal behavior. Mice heterozygous for a disrupted allele of the homologous Taok3 gene (Taok3Gt) were resistant to the acute sedative effects of ethanol. JNK activity was constitutively increased in brains of Taok3Gt/+ mice, and acute induction of phospho-JNK in brain tissue by ethanol was occluded in Taok3Gt/+ mice. Finally, acute administration of a JNK inhibitor conferred resistance to the sedative effects of ethanol in wild-type but not Taok3Gt/+ mice. Taken together, these data support a role of a TAO/TAOK3-JNK neuronal signaling pathway in regulating sensitivity to acute ethanol exposure in flies and in mice.
Sexual behaviors in animals are governed by inputs from multiple external sensory modalities. However, how these inputs are integrated to jointly control animal behavior is still poorly understood. Whereas visual information alone is not sufficient to induce courtship behavior in Drosophila melanogaster males, when a subset of male-specific fruitless (fru)- and doublesex (dsx)-expressing neurons that respond to chemosensory cues (P1 neurons) were artificially activated via a temperature-sensitive cation channel (dTRPA1), males followed and extended their wing toward moving objects (even a moving piece of rubber band) intensively. When stationary, these objects were not courted. Our results indicate that motion input and activation of P1 neurons are individually necessary, and under our assay conditions, jointly sufficient to elicit early courtship behaviors, and provide insights into how courtship decisions are made via sensory integration.
Large electron microscopy image datasets for connectomics are typically composed of thousands to millions of partially overlapping two-dimensional images (tiles), which must be registered into a coherent volume prior to further analysis. A common registration strategy is to find matching features between neighboring and overlapping image pairs, followed by a numerical estimation of optimal image deformation using a so-called solver program.
Existing solvers are inadequate for large data volumes, and inefficient for small-scale image registration.
In this work, an efficient and accurate matrix-based solver method is presented. A linear system is constructed that combines minimization of feature-pair square distances with explicit constraints in a regularization term. In absence of reliable priors for regularization, we show how to construct a rigid-model approximation to use as prior. The linear system is solved using available computer programs, whose performance on typical registration tasks we briefly compare, and to which future scale-up is delegated. Our method is applied to the joint alignment of 2.67 million images, with more than 200 million point-pairs and has been used for successfully aligning the first full adult fruit fly brain.
The correction of bias in magnetic resonance images is an important problem in medical image processing. Most previous approaches have used a maximum likelihood method to increase the likelihood of the pixels in a single image by adaptively estimating a correction to the unknown image bias field. The pixel likelihoods are defined either in terms of a pre-existing tissue model, or non-parametrically in terms of the image’s own pixel values. In both cases, the specific location of a pixel in the image is not used to calculate the likelihoods. We suggest a new approach in which we simultaneously eliminate the bias from a set of images of the same anatomy, but from different patients. We use the statistics from the same location across different images, rather than within an image, to eliminate bias fields from all of the images simultaneously. The method builds a multi-resolution non-parametric tissue model conditioned on image location while eliminating the bias fields associated with the original image set. We present experiments on both synthetic and real MR data sets, and present comparisons with other methods.
To compare spatial patterns of gene expression, one must analyze a large number of images as current methods are only able to measure a small number of genes at a time. Bringing images of corresponding tissues into alignment is a critical first step in making a meaningful comparative analysis of these spatial patterns. Significant image noise and variability in the shapes make it hard to pick a canonical shape model. In this paper, we address these problems by combining segmentation and unsupervised shape learning algorithms. We first segment images to acquire structures of interest, then jointly align the shapes of these acquired structures using an unsupervised nonparametric maximum likelihood algorithm along the lines of congealing, while simultaneously learning the underlying shape model and associated transformations. The learned transformations are applied to corresponding images to bring them into alignment in one step. We demonstrate the results for images of various classes of Drosophila imaginal discs and discuss the methodology used for a quantitative analysis of spatial gene expression patterns.
Infective juveniles of the insect-parastic nematode canjump greater than 6 times their height, a striking evolved novelty in some species of this genus. Using high-speed videography, we observed the kinematics of spontaneousjumping behavior. Our analysis places a lower bound on the velocity and acceleration of these worms.