Filter
Associated Lab
- Aguilera Castrejon Lab (4) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (64) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (104) Apply Betzig Lab filter
- Beyene Lab (10) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (53) Apply Branson Lab filter
- Card Lab (37) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (15) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dennis Lab (2) Apply Dennis Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (46) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (21) Apply Espinosa Medina Lab filter
- Feliciano Lab (16) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (17) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (55) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (28) Apply Hermundstad Lab filter
- Hess Lab (77) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (45) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (2) Apply Johnson Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (62) Apply Keller Lab filter
- Koay Lab (4) Apply Koay Lab filter
- Lavis Lab (150) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (8) Apply Li Lab filter
- Lippincott-Schwartz Lab (110) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (4) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (60) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (7) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (44) Apply Pachitariu Lab filter
- Pastalkova Lab (6) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (49) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (40) Apply Romani Lab filter
- Rubin Lab (111) Apply Rubin Lab filter
- Saalfeld Lab (49) Apply Saalfeld Lab filter
- Satou Lab (3) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (55) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (3) Apply Sgro Lab filter
- Shroff Lab (35) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (62) Apply Spruston Lab filter
- Stern Lab (77) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (41) Apply Stringer Lab filter
- Svoboda Lab (132) Apply Svoboda Lab filter
- Tebo Lab (12) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (19) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (41) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (5) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (11) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (49) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (18) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- High Performance Computing (7) Apply High Performance Computing filter
- Integrative Imaging (25) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (40) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Mass Spectrometry (1) Apply Mass Spectrometry filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Project Technical Resources (54) Apply Project Technical Resources filter
- Quantitative Genomics (20) Apply Quantitative Genomics filter
- Scientific Computing (106) Apply Scientific Computing filter
- Stem Cell & Primary Culture (14) Apply Stem Cell & Primary Culture filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2026 (118) Apply 2026 filter
- 2025 (223) Apply 2025 filter
- 2024 (208) Apply 2024 filter
- 2023 (157) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2896 Janelia Publications
Showing 1411-1420 of 2896 resultsThe ability to discriminate sensory stimuli with overlapping features is thought to arise in brain structures called expansion layers, where neurons carrying information about sensory features make combinatorial connections onto a much larger set of cells. For 50 years, expansion coding has been a prime topic of theoretical neuroscience, which seeks to explain how quantitative parameters of the expansion circuit influence sensory sensitivity, discrimination, and generalization. Here, we investigate the developmental events that produce the quantitative parameters of the arthropod expansion layer, called the mushroom body. Using Drosophila melanogaster as a model, we employ genetic and chemical tools to engineer changes to circuit development. These allow us to produce living animals with hypothesis-driven variations on natural expansion layer wiring parameters. We then test the functional and behavioral consequences. By altering the number of expansion layer neurons (Kenyon cells) and their dendritic complexity, we find that input density, but not cell number, tunes neuronal odor selectivity. Simple odor discrimination behavior is maintained when the Kenyon cell number is reduced and augmented by Kenyon cell number expansion. Animals with increased input density to each Kenyon cell show increased overlap in Kenyon cell odor responses and become worse at odor discrimination tasks.
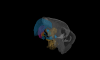
The basal ganglia play a critical role in the regulation of voluntary action in vertebrates. Our understanding of the function of the basal ganglia relies heavily upon anatomical information, but continued progress will require an understanding of the specific functional roles played by diverse cell types and their connectivity. An increasing number of mouse lines allow extensive identification, characterization, and manipulation of specified cell types in the basal ganglia. Despite the promise of genetically modified mice for elucidating the functional roles of diverse cell types, there is relatively little anatomical data obtained directly in the mouse. Here we have characterized the retrograde labeling obtained from a series of tracer injections throughout the dorsal striatum of adult mice. We found systematic variations in input along both the medial-lateral and anterior-posterior neuraxes in close agreement with canonical features of basal ganglia anatomy in the rat. In addition to the canonical features we have provided experimental support for the importance of non-canonical inputs to the striatum from the raphe nuclei and the amygdala. To look for organization at a finer scale we have analyzed the correlation structure of labeling intensity across our entire dataset. Using this analysis we found substantial local heterogeneity within the large-scale order. From this analysis we conclude that individual striatal sites receive varied combinations of cortical and thalamic input from multiple functional areas, consistent with some earlier studies in the rat that have suggested the presence of a combinatorial map.
Fruit flies (Drosophila melanogaster) are small insects, with correspondingly small power budgets. Despite this, they perform sophisticated neural computations in real time. Careful study of these insects is revealing how some of these circuits work. Insights from these systems might be helpful in designing other low power circuits.
The mammalian vomeronasal organ encodes pheromone information about gender, reproductive status, genetic background and individual differences. It remains unknown how pheromone information interacts to trigger innate behaviors. In this study, we identify vomeronasal receptors responsible for detecting female pheromones. A sub-group of V1re clade members recognizes gender-identifying cues in female urine. Multiple members of the V1rj clade are cognate receptors for urinary estrus signals, as well as for sulfated estrogen (SE) compounds. In both cases, the same cue activates multiple homologous receptors, suggesting redundancy in encoding female pheromone cues. Neither gender-specific cues nor SEs alone are sufficient to promote courtship behavior in male mice, whereas robust courtship behavior can be induced when the two cues are applied together. Thus, integrated action of different female cues is required in pheromone-triggered mating behavior. These results suggest a gating mechanism in the vomeronasal circuit in promoting specific innate behavior.DOI: http://dx.doi.org/10.7554/eLife.03025.001.
The nervous system evolved to enable navigation throughout the environment in the pursuit of resources. Evolutionarily newer structures allowed increasingly complex adaptations but necessarily added redundancy. A dominant view of movement neuroscientists is that there is a one-to-one mapping between brain region and function. However, recent experimental data is hard to reconcile with the most conservative interpretation of this framework, suggesting a degree of functional redundancy during the performance of well-learned, constrained behaviors. This apparent redundancy likely stems from the bidirectional interactions between the various cortical and subcortical structures involved in motor control. We posit that these bidirectional connections enable flexible interactions across structures that change depending upon behavioral demands, such as during acquisition, execution or adaptation of a skill. Observing the system across both multiple actions and behavioral timescales can help isolate the functional contributions of individual structures, leading to an integrated understanding of the neural control of movement.
Elucidating the diversity and spatial organization of cell types in the brain is an essential goal of neuroscience, with many emerging technologies helping to advance this endeavor. Using a new in situ hybridization method that can measure the expression of hundreds of genes in a given mouse brain section (amplified seqFISH), Shah et al. (2016) describe a spatial organization of hippocampal cell types that differs from previous reports. In seeking to understand this discrepancy, we find that many of the barcoded genes used by seqFISH to characterize this spatial organization, when cross-validated by other sensitive methodologies, exhibit negligible expression in the hippocampus. Additionally, the results of Shah et al. (2016) do not recapitulate canonical cellular hierarchies and improperly classify major neuronal cell types. We suggest that, when describing the spatial organization of brain regions, cross-validation using multiple techniques should be used to yield robust and informative cellular classification. This Matters Arising paper is in response to Shah et al. (2016), published in Neuron. See also the response by Shah et al. (2017), published in this issue.
An understanding of human brain individuality requires the integration of data on brain organization across people and brain regions, molecular and systems scales, as well as healthy and clinical states. Here, we help advance this understanding by leveraging methods from computational genomics to integrate large-scale genomic, transcriptomic, neuroimaging, and electronic-health record data sets. We estimated genetically regulated gene expression (gr-expression) of 18,647 genes, across 10 cortical and subcortical regions of 45,549 people from the UK Biobank. First, we showed that patterns of estimated gr-expression reflect known genetic-ancestry relationships, regional identities, as well as inter-regional correlation structure of directly assayed gene expression. Second, we performed transcriptome-wide association studies (TWAS) to discover 1,065 associations between individual variation in gr-expression and gray-matter volumes across people and brain regions. We benchmarked these associations against results from genome-wide association studies (GWAS) of the same sample and found hundreds of novel associations relative to these GWAS. Third, we integrated our results with clinical associations of gr-expression from the Vanderbilt Biobank. This integration allowed us to link genes, via gr-expression, to neuroimaging and clinical phenotypes. Fourth, we identified associations of polygenic gr-expression with structural and functional MRI phenotypes in the Human Connectome Project (HCP), a small neuroimaging-genomic data set with high-quality functional imaging data. Finally, we showed that estimates of gr-expression and magnitudes of TWAS were generally replicable and that the p-values of TWAS were replicable in large samples. Collectively, our results provide a powerful new resource for integrating gr-expression with population genetics of brain organization and disease.
Accurately predicting an outcome requires that animals learn supporting and conflicting evidence from sequential experience. In mammals and invertebrates, learned fear responses can be suppressed by experiencing predictive cues without punishment, a process called memory extinction. Here, we show that extinction of aversive memories in Drosophila requires specific dopaminergic neurons, which indicate that omission of punishment is remembered as a positive experience. Functional imaging revealed co-existence of intracellular calcium traces in different places in the mushroom body output neuron network for both the original aversive memory and a new appetitive extinction memory. Light and ultrastructural anatomy are consistent with parallel competing memories being combined within mushroom body output neurons that direct avoidance. Indeed, extinction-evoked plasticity in a pair of these neurons neutralizes the potentiated odor response imposed in the network by aversive learning. Therefore, flies track the accuracy of learned expectations by accumulating and integrating memories of conflicting events.
Nuclear pore complexes play central roles as gatekeepers of RNA and protein transport between the cytoplasm and nucleoplasm. However, their large size and dynamic nature have impeded a full structural and functional elucidation. Here we determined the structure of the entire 552-protein nuclear pore complex of the yeast Saccharomyces cerevisiae at sub-nanometre precision by satisfying a wide range of data relating to the molecular arrangement of its constituents. The nuclear pore complex incorporates sturdy diagonal columns and connector cables attached to these columns, imbuing the structure with strength and flexibility. These cables also tie together all other elements of the nuclear pore complex, including membrane-interacting regions, outer rings and RNA-processing platforms. Inwardly directed anchors create a high density of transport factor-docking Phe-Gly repeats in the central channel, organized into distinct functional units. This integrative structure enables us to rationalize the architecture, transport mechanism and evolutionary origins of the nuclear pore complex.
Due to their small size and transparency, zebrafish larvae are amenable to a range of fluorescence microscopy techniques. With the development of sensitive genetically encoded calcium indicators, this has extended to the whole-brain imaging of neural activity with cellular resolution. This technique has been used to study brain-wide population dynamics accompanying sensory processing and sensorimotor transformations, and has spurred the development of innovative closed-loop behavioral paradigms in which stimulus-response relationships can be studied. More recently, microscopes have been developed that allow whole-brain calcium imaging in freely swimming and behaving larvae. In this review, we highlight the technologies underlying whole-brain functional imaging in zebrafish, provide examples of the sensory and motor processes that have been studied with this technique, and discuss the need to merge data from whole-brain functional imaging studies with neurochemical and anatomical information to develop holistic models of functional neural circuits.