Filter
Associated Lab
- Aguilera Castrejon Lab (2) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (58) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (103) Apply Betzig Lab filter
- Beyene Lab (10) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (51) Apply Branson Lab filter
- Card Lab (37) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (41) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (18) Apply Espinosa Medina Lab filter
- Feliciano Lab (10) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (42) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (55) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (26) Apply Hermundstad Lab filter
- Hess Lab (76) Apply Hess Lab filter
- Ilanges Lab (3) Apply Ilanges Lab filter
- Jayaraman Lab (44) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (3) Apply Koay Lab filter
- Lavis Lab (144) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (6) Apply Li Lab filter
- Lippincott-Schwartz Lab (107) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (2) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (59) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (39) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (49) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (39) Apply Romani Lab filter
- Rubin Lab (110) Apply Rubin Lab filter
- Saalfeld Lab (47) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (52) Apply Schreiter Lab filter
- Sgro Lab (2) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (61) Apply Spruston Lab filter
- Stern Lab (75) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (36) Apply Stringer Lab filter
- Svoboda Lab (132) Apply Svoboda Lab filter
- Tebo Lab (11) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (18) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (41) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (4) Apply Voigts Lab filter
- Wang (Meng) Lab (27) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (9) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (41) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (18) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- High Performance Computing (7) Apply High Performance Computing filter
- Integrative Imaging (18) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (37) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Mass Spectrometry (1) Apply Mass Spectrometry filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (14) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (53) Apply Project Technical Resources filter
- Quantitative Genomics (20) Apply Quantitative Genomics filter
- Scientific Computing (100) Apply Scientific Computing filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2026 (17) Apply 2026 filter
- 2025 (225) Apply 2025 filter
- 2024 (211) Apply 2024 filter
- 2023 (157) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2800 Janelia Publications
Showing 691-700 of 2800 resultsDuring postembryonic development, the nervous system must adapt to a growing body. How changes in neuronal structure and connectivity contribute to the maintenance of appropriate circuit function remains unclear. In a previous paper (Schneider-Mizell et al., 2016), we measured the cellular neuroanatomy underlying synaptic connectivity in Drosophila. Here, we examined how neuronal morphology and connectivity change between 1st instar and 3rd instar larval stages using serial section electron microscopy. We reconstructed nociceptive circuits in a larva of each stage and found consistent topographically arranged connectivity between identified neurons. Five-fold increases in each size, number of terminal dendritic branches, and total number of synaptic inputs were accompanied by cell-type specific connectivity changes that preserved the fraction of total synaptic input associated with each presynaptic partner. We propose that precise patterns of structural growth act to conserve the computational function of a circuit, for example determining the location of a dangerous stimulus.
Numerous efforts to generate "connectomes," or synaptic wiring diagrams, of large neural circuits or entire nervous systems are currently underway. These efforts promise an abundance of data to guide theoretical models of neural computation and test their predictions. However, there is not yet a standard set of tools for incorporating the connectivity constraints that these datasets provide into the models typically studied in theoretical neuroscience. This article surveys recent approaches to building models with constrained wiring diagrams and the insights they have provided. It also describes challenges and the need for new techniques to scale these approaches to ever more complex datasets.
"Regulatory evolution," that is, changes in a gene's expression pattern through changes at its regulatory sequence, rather than changes at the coding sequence of the gene or changes of the upstream transcription factors, has been increasingly recognized as a pervasive evolution mechanism. Many somatic sexually dimorphic features of Drosophila melanogaster are the results of gene expression regulated by the doublesex (dsx) gene, which encodes sex-specific transcription factors (DSX(F) in females and DSX(M) in males). Rapid changes in such sexually dimorphic features are likely a result of changes at the regulatory sequence of the target genes. We focused on the Flavin-containing monooxygenase-2 (Fmo-2) gene, a likely direct dsx target, to elucidate how sexually dimorphic expression and its evolution are brought about. We found that dsx is deployed to regulate the Fmo-2 transcription both in the midgut and in fat body cells of the spermatheca (a female-specific tissue), through a canonical DSX-binding site in the Fmo-2 regulatory sequence. In the melanogaster group, Fmo-2 transcription in the midgut has evolved rapidly, in contrast to the conserved spermathecal transcription. We identified two cis-regulatory modules (CRM-p and CRM-d) that direct sexually monomorphic or dimorphic Fmo-2 transcription, respectively, in the midguts of these species. Changes of Fmo-2 transcription in the midgut from sexually dimorphic to sexually monomorphic in some species are caused by the loss of CRM-d function, but not the loss of the canonical DSX-binding site. Thus, conferring transcriptional regulation on a CRM level allows the regulation to evolve rapidly in one tissue while evading evolutionary constraints posed by other tissues.
Close appositions between the membrane of the endoplasmic reticulum (ER) and other intracellular membranes have important functions in cell physiology. These include lipid homeostasis, regulation of Ca(2+) dynamics, and control of organelle biogenesis and dynamics. Although these membrane contacts have previously been observed in neurons, their distribution and abundance have not been systematically analyzed. Here, we have used focused ion beam-scanning electron microscopy to generate 3D reconstructions of intracellular organelles and their membrane appositions involving the ER (distance ≤30 nm) in different neuronal compartments. ER-plasma membrane (PM) contacts were particularly abundant in cell bodies, with large, flat ER cisternae apposed to the PM, sometimes with a notably narrow lumen (thin ER). Smaller ER-PM contacts occurred throughout dendrites, axons, and in axon terminals. ER contacts with mitochondria were abundant in all compartments, with the ER often forming a network that embraced mitochondria. Small focal contacts were also observed with tubulovesicular structures, likely to be endosomes, and with sparse multivesicular bodies and lysosomes found in our reconstructions. Our study provides an anatomical reference for interpreting information about interorganelle communication in neurons emerging from functional and biochemical studies.
The representation of contextual information peripheral to a salient stimulus is central to an animal's ability to correctly interpret and flexibly respond to that stimulus. While the computations and circuits underlying the context-dependent modulation of stimulus-response pairings have typically been studied in vertebrates, the genetic tractability, numeric simplification, and well-characterized connectivity patterns of the Drosophila melanogaster brain have facilitated circuit-level insights into contextual processing. Recent studies in flies reveal the neuronal mechanisms that create flexible context-dependent behavioral responses to sensory events in conditions of predation threat, feeding regulation, and social interaction.
The brain is an organ of immense complexity. Next-generation RNA sequencing (RNA-seq) is becoming increasingly popular in the deconstruction of this complexity into distinct classes of 'cell types'. Notably, in addition to revealing the organization of this distinct cell-type landscape, the technology has also begun to illustrate that continuous variation can be found within narrowly defined cell types. Here we summarize the evidence for graded transcriptomic heterogeneity being present, widespread, and functionally relevant in the nervous system. We explain how these graded differences can map onto higher-order organizational features and how they may reframe existing interpretations of higher-order heterogeneity. Ultimately, a multimodal approach incorporating continuously variable cell types will facilitate an accurate reductionist interpretation of the nervous system.
In vivo imaging at high spatiotemporal resolution is key to the understanding of complex biological systems. We integrated an optical phase-locked ultrasound lens into a two-photon fluorescence microscope and achieved microsecond-scale axial scanning, thus enabling volumetric imaging at tens of hertz. We applied this system to multicolor volumetric imaging of processes sensitive to motion artifacts, including calcium dynamics in behaving mouse brain and transient morphology changes and trafficking of immune cells.
Mechano-transduction is an emerging but still poorly understood component of T cell activation. Here we investigated the ligand-dependent contribution made by contractile actomyosin arcs populating the peripheral supramolecular activation cluster (pSMAC) region of the immunological synapse (IS) to T cell receptor (TCR) microcluster transport and proximal signaling in primary mouse T cells. Using super resolution microscopy, OT1-CD8+ mouse T cells, and two ovalbumin (OVA) peptides with different affinities for the TCR, we show that the generation of organized actomyosin arcs depends on ligand potency and the ability of myosin 2 to contract actin filaments. While weak ligands induce disorganized actomyosin arcs, strong ligands result in organized actomyosin arcs that correlate well with tension-sensitive CasL phosphorylation and the accumulation of ligands at the IS center. Blocking myosin 2 contractility greatly reduces the difference in the extent of Src and LAT phosphorylation observed between the strong and the weak ligand, arguing that myosin 2-dependent force generation within actin arcs contributes to ligand discrimination. Together, our data are consistent with the idea that actomyosin arcs in the pSMAC region of the IS promote a mechano-chemical feedback mechanism that amplifies the accumulation of critical signaling molecules at the IS.
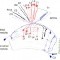
Motion detection is a fundamental neural computation performed by many sensory systems. In the fly, local motion computation is thought to occur within the first two layers of the visual system, the lamina and medulla. We constructed specific genetic driver lines for each of the 12 neuron classes in the lamina. We then depolarized and hyperpolarized each neuron type and quantified fly behavioral responses to a diverse set of motion stimuli. We found that only a small number of lamina output neurons are essential for motion detection, while most neurons serve to sculpt and enhance these feedforward pathways. Two classes of feedback neurons (C2 and C3), and lamina output neurons (L2 and L4), are required for normal detection of directional motion stimuli. Our results reveal a prominent role for feedback and lateral interactions in motion processing and demonstrate that motion-dependent behaviors rely on contributions from nearly all lamina neuron classes.
The Drosophila mushroom body (MB) is an associative learning network that is important for the control of sleep. We have recently identified particular intrinsic MB Kenyon cell (KC) classes that regulate sleep through synaptic activation of particular MB output neurons (MBONs) whose axons convey sleep control signals out of the MB to downstream target regions. Specifically, we found that sleep-promoting KCs increase sleep by preferentially activating cholinergic sleep-promoting MBONs, while wake-promoting KCs decrease sleep by preferentially activating glutamatergic wake-promoting MBONs. Here we use a combination of genetic and physiological approaches to identify wake-promoting dopaminergic neurons (DANs) that innervate the MB, and show that they activate wake-promoting MBONs. These studies reveal a dopaminergic sleep control mechanism that likely operates by modulation of KC-MBON microcircuits.